




2. 泛亚太生物科技有限公司, 东莞 523808
2. Asiapac Biotechnology Co., Ltd., Dongguan 523808, China
果胶是存在于植物细胞壁和细胞间的一种黏连物质,它是半乳糖醛酸以α-1,4键连接的[1]。在成熟的植物细胞壁中,纤维素、半纤维素、木质素和果胶紧密结合在一起,因而大大降低了动物对细胞壁物质的可消化性,而动物肠道内缺乏分解木质素和果胶的酶,同时由于果胶对纤维素、半纤维素的包裹和黏连作用,也阻碍了某些酶对纤维素和半纤维的作用,只有在果胶分解酶的作用下,分解果胶后才能释放出纤维素和半纤维素[2]。目前酶制剂的研究多集中在复合酶制剂,对单体酶的研究主要以纤维素酶、植酸酶和淀粉酶为主,对饲用果胶酶并没有进行深入的研究[3]。复合酶制剂针对的底物较多,在动物生产上的作用一般要大于单体酶,但单体酶作用效果和作用机理的研究会对复合酶制剂的研究起重要的指导作用。果胶酶作为非淀粉多糖酶的一种,有许多重要功能,如降解植物细胞壁、消除抗营养因子、改善肠道功能、提高内源酶活性等[3]。因此,对果胶酶的性质、饲料中的适宜添加量以及与其他酶的复合协同作用等方面的研究具有重要意义。本试验测定了肉鸡对几种常规和非常规饲料原料的回肠氨基酸消化率和代谢能,并研究了添加单一成分果胶酶后对其的改进效果,旨在为丰富这些原料的数据库以及果胶酶的使用提供参考依据。
本试验所用豆粕分为优质豆粕(HQSBM,粗蛋白质44.77%)和普通豆粕(SBM,粗蛋白质42.54%);菜籽粕为普通菜籽粕(RSM);棉籽粕分为普通棉籽粕(CSM)、脱酚棉籽粕(PCSM)和棉籽蛋白(CSP),试验原料均由河北大午集团提供。果胶酶由泛亚太(东莞)生物科技有限公司提供,酶活性为9 700 U/g。
试验选用780只1日龄罗斯308肉公鸡,在非试验期饲喂符合我国《鸡饲养标准》(NY/T 33—200)的全价配合饲粮,基础饲粮组成及营养水平见表1。在15日龄对所有肉鸡根据体重重新分为13个处理(每个处理6个重复,每个重复10只鸡),其中12个处理分别饲喂豆粕、棉籽粕和菜籽粕型半纯合饲粮,并分别设添加果胶酶组与空白组;另外1个处理饲喂无氮饲粮以测定氨基酸内源损失。16~18日龄开始预饲试验饲粮,以待测饲料原料作为唯一的饲粮蛋白质来源,粗蛋白质水平为20%左右,尽量保证除氨基酸外的营养平衡。19~23日龄为正式试验期。试验饲粮的果胶酶添加量均为100 mg/kg,不添加酶组为空白对照。无氮饲粮组成见表2,6种饲料原料的常规营养成分实测值见表3,试验饲粮组成见表4,试验饲粮粗蛋白质及氨基酸含量实测值见表5。试验用外源指示剂为二氧化钛,添加量为0.5%。整个试验阶段均保证充足的饮水、自由采食及正常的免疫程序。
| 表1 基础饲粮组成及营养水平(风干基础) Table 1 Composition and nutrient levels of the basal diet (air-dry basis) % |
| 表2 无氮饲粮组成(风干基础) Table 2 Composition of the nitrogen-free diet (air-dry basis) % |
| 表3 6种饲料原料的常规营养成分实测值 Table 3 Measured values of conventional nutrients of six kinds of feedstuffs % |
| 表4 试验饲粮组成(风干基础)
Table 4 Composition of experimental diets (air-dry basis) %
|
| 表5 试验饲粮粗蛋白质及氨基酸含量实测值(风干基础)
Table 5 Measured values of crude protein and amino acid contents of experimental diets (air-dry basis) %
|
肉鸡从19日龄开始禁食24 h,禁食结束后安装收粪盘,20~21日龄饲喂试验饲粮并且记录2 d的采食量,22日龄继续禁食24 h,收粪期为3 d(20~22日龄),收粪以1个重复为1个单位,每天收集的粪便用塑料袋装好称重后放入-20 ℃冰箱保存,收集完所有粪便后解冻,利用鼓风干燥箱在65 ℃恒温进行烘干处理,粉碎后过40目筛,用大培养皿取400 g左右,再进一步进行指标测定。
肉鸡在23日龄开始自由采食,采集食糜前先饥饿4 h,采食1 h,再过3 h后开始宰杀取样。从每个处理各个重复挑选10只鸡,翅静脉戊巴比妥钠(30 mg/kg)麻醉后,立即剖腹,分离整段回肠。回肠食糜取后半段(弃去最后2 cm),吸取少量蒸馏水冲出回肠食糜于铝盒中。2种食糜均先于-20 ℃冰箱保存,之后采用冷冻干燥机冻干食糜后备测。
测定方法参照《饲料分析鸡饲料质量检测技术》(第3版)。
胱氨酸和蛋氨酸含量的测定采用过甲酸氧化再用高效液相色谱(HPLC)法测定;色氨酸用氢氧化钡水解后再用HPLC法测定,其余氨基酸含量用6 mol/L盐酸水解后用氨基酸自动分析仪(日立835-50)进行测定,检测单位为农业部饲料效价与安全监督检验测试中心(北京),具体方法参照GB/T 18246—2000。
式中:AME葡萄糖为12.89 MJ/kg;FE为饲料总能(MJ/kg);EE为粪便总能(MJ/kg);FI为总的采食量(kg);EO为粪便排出量(kg)。所有含量都以干物质为基础。
AMEn豆粕/菜籽粕/棉籽粕(MJ/kg)=AME豆粕/菜籽粕/棉籽粕-RN×34.39。式中:RN为家禽每日采食每千克饲粮所沉积的氮量(g);34.39为沉积每克氮需要的能量(kJ)。
式中:AAi为回肠食糜氨基酸含量;TDCi为回肠食糜二氧化钛含量;AAd为饲粮氨基酸含量;TDCd为饲粮二氧化钛含量。所有含量都以干物质为基础。
式中:IAAend为对应每单位重量的回肠末端内源氨基酸损失的某氨基酸量(惯用表示为mg/kg);AAi为无氮饲粮回肠食糜某氨基酸含量(冻干基础);TDCd、TDCi分别为无氮饲粮二氧化钛含量(干物质基础)、回肠食糜二氧化钛含量(冻干基础)。
式中:AAd为待测饲粮干物质状态下某氨基酸含量。
采用SPSS 17.0统计软件中一般线性模型中多变量对豆粕进行2×2双因素方差分析(饲料原料×酶),对棉籽粕进行3×2双因素方差分析(饲料原料×酶),对菜籽粕进行单因素(酶)方差分析,以P≤0.05作为差异显著性标准。
由表6可知,优质豆粕和普通豆粕的AME和AMEn均存在极显著差异(P<0.01),且优质豆粕高于普通豆粕;添加果胶酶后,2种豆粕的AME和AMEn均存在提高趋势(P=0.066),优质豆粕和普通豆粕的AME提高幅度分别为2.2%、8.1%,AMEn的提高幅度分别为2.1%、8.1%;原料和酶之间不存在互作效应(P>0.05)。
| 表6 果胶酶对2种豆粕AME和AMEn的影响(干物质基础)
Table 6 Effects of pectinase on AME and AMEn of two kinds of soybean meals (DM basis) MJ/kg
|
由表7可知,3种棉籽粕的AME和AMEn存在显著差异(P<0.05),其表现趋势为棉籽蛋白最高,脱酚棉籽粕其次,普通棉籽粕最低;添加果胶酶后,3种棉籽粕的AME和AMEn均得到极显著提高(P<0.01),AME中棉籽蛋白提高幅度最大,达7.7%,其次是普通棉籽粕,为7.3%,脱酚棉籽粕的提高幅度最少,为2.2%,AMEn的提高幅度则分别为7.4%、6.6%和1.8%;原料和酶之间不存在互作效应(P>0.05)。
| 表7 果胶酶对3种棉籽粕AME和AMEn的影响(干物质基础)
Table 7 Effects of pectinase on AME and AMEn of three kinds of cottonseed meals (DM basis) MJ/kg
|
由表8可知,菜籽粕在添加果胶酶后,AME和AMEn虽然变化不显著(P>0.05),但均有所提高,提高幅度分别为2.5%、2.0%。
| 表8 果胶酶对菜籽粕AME和AMEn的影响(干物质基础) Table 8 Effects of pectinase on AME and AMEn of two kinds of rapeseed meal (DM basis) MJ/kg |
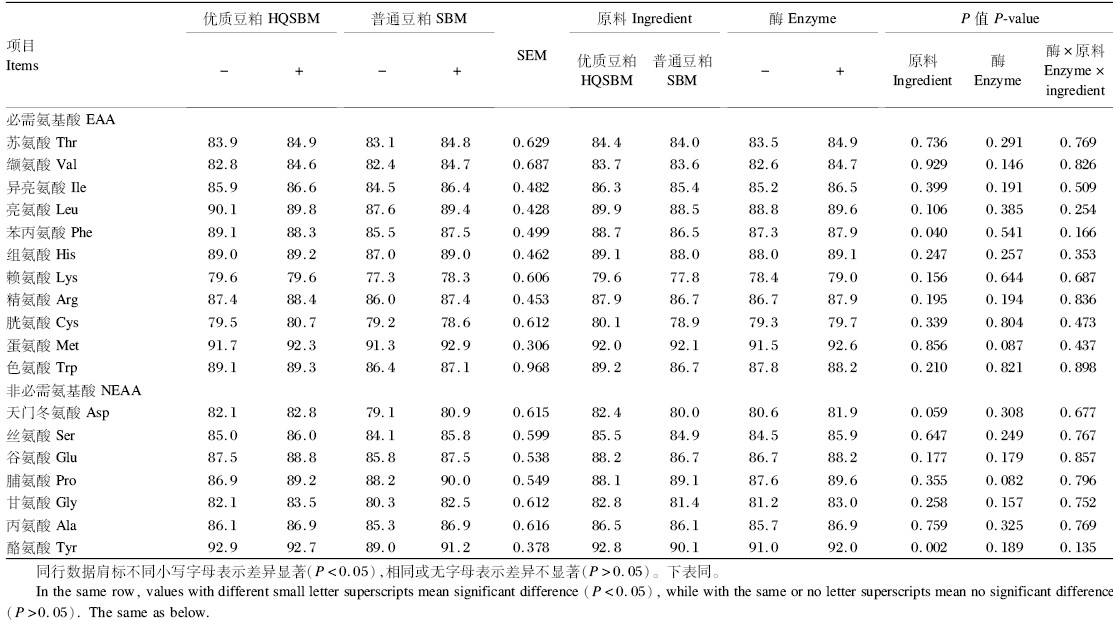
由表9可知,肉鸡对优质豆粕中苯丙氨酸和酪氨酸的SID显著高于普通豆粕(P<0.05),天门冬氨酸则表现出差异趋势(P=0.059),其余氨基酸的SID在2种豆粕间不存在显著差异(P>0.05),但从数值上可以看出优质豆粕表现出一定优势;添加果胶酶后,只有蛋氨酸和脯氨酸的SID表现出提高趋势(P=0.087、P=0.082),其余氨基酸均无显著差异(P>0.05),但从数值上可以看出,添加果胶酶后所有氨基酸的SID均有一定程度的提高;氨基酸消化率在原料和酶之间并不存在互作效应(P>0.05)。
| 表9 果胶酶对2种豆粕中氨基酸标准回肠消化率的影响 Table9 Effects of pectinase on AA SID of two kinds of soybean meal |
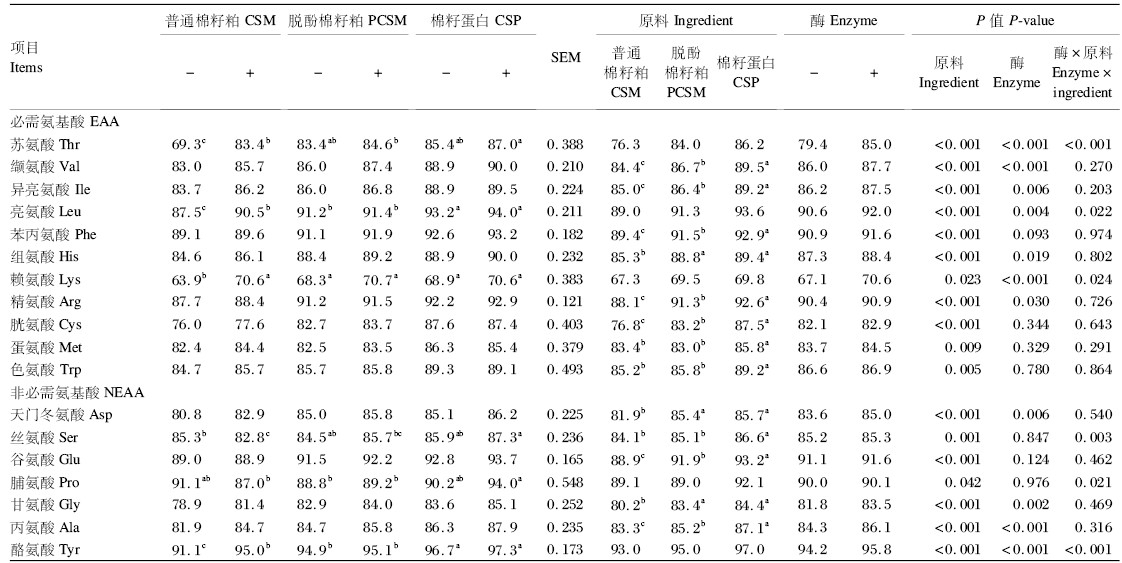
由表10可知,3种棉籽粕中所有氨基酸的SID均存在显著差异(P<0.05),其氨基酸的SID表现规律为棉籽蛋白高于脱酚棉籽粕高于普通棉籽粕;添加果胶酶后,除了苯丙氨酸、胱氨酸、蛋氨酸、色氨酸、丝氨酸、谷氨酸和脯氨酸外,其余氨基酸的SID均显著提高(P<0.05),表明果胶酶对棉籽粕的氨基酸的SID具有较大的改善作用;苏氨酸、亮氨酸、赖氨酸、丝氨酸、脯氨酸和酪氨酸的SID在原料和酶之间存在互作效应(P<0.05),其余氨基酸的SID则不存在互作效应(P>0.05)。
| 表10 果胶酶对3种棉籽粕中氨基酸标准回肠消化率的影响 Table10 Effects of pectinase on AA SID of three kinds of cottonseed meal |
由表11可知,菜籽粕在添加果胶酶后,大部分氨基酸的SID提高趋势不显著(P>0.05),但在数值上仍有提高;但酪氨酸的SID有降低趋势(P=0.073),亮氨酸、苯丙氨酸、色氨酸、精氨酸和天门冬氨酸的SID则在数值上略有降低,表明添加果胶酶后肉鸡对菜籽粕的氨基酸的SID没有显著改善作用。
| 表11 果胶酶对菜籽粕中氨基酸标准回肠消化率的影响 Table 11 Effects of pectinase on AA SID of rapeseed meal % |
测定方法、试验动物和饲料本身是影响家禽饲料代谢能的3个主要因素[6, 7]。Mateos等[8]研究表明,饲粮油脂的添加水平、脂肪酸组成均会对代谢能产生影响,油脂在一定范围内随着添加量的增加,代谢能会提高。但油脂的添加量增加到一定范围时,代谢能不再增加反而会下降[9],这说明脂肪在一定程度上影响着饲料的代谢能值。另外,Villamide等[10]研究发现,饲粮纤维素对饲料代谢能值的影响非常大,其含量与饲料代谢能呈高度负相关。郑萍等[11]和席鹏彬等[12]研究也发现,饲粮中性纤维水平与其营养物质消化率有着负相关关系。
由上可知,在本试验中影响肉鸡代谢能的主要因素是饲料,优质豆粕中粗脂肪含量高于普通豆粕,而粗纤维含量低于普通豆粕,这可能是导致其代谢能和氨基酸消化率高的主要原因。虽然普通棉籽粕中粗脂肪含量高于脱酚棉籽粕和棉籽蛋白,但普通棉籽粕的纤维含量大幅高于脱酚棉籽粕和棉籽蛋白,脂肪对代谢能的贡献不足以抵消过高粗纤维带来的负面效应,因而其代谢能和氨基酸消化率最低,脱酚棉籽粕居中,而棉籽蛋白最高。
另外,饲料中多种抗营养因子均会影响营养物质的消化吸收与利用[13, 14, 15],豆粕、棉籽粕和菜籽粕中均含大量的抗营养因子。
本试验中优质豆粕的抗营养因子少于普通豆粕;棉籽蛋白和脱酚棉籽粕经过特殊工艺处理,其抗营养因子数量大幅减少,而普通棉籽粕由于没有经过处理,含有大量的棉酚、环丙烯脂肪酸、植酸、单宁以及非淀粉多糖[16],这也是导致这些原料代谢能和氨基酸消化率出现差异的另一个原因。
果胶是存在于所有高等植物初生壁和细胞间隙中一种密切相关的多糖,是非淀粉多糖中化学组分和分子结果变化最大的一类多糖,在不同植物组织中其化学结构和相对分子质量变化很大[1]。果胶在初生壁中与不同含量的纤维素、半纤维素、木质素的微纤丝以及某些伸展蛋白相互交联,使各种细胞组织结构坚硬,表现出固有的形态[15],因此极大地阻碍了细胞壁营养物质的吸收。Schneeman等[16]研究发现,果胶能够在肠道产生黏胶,增加食糜黏度;Roberfroid等[17]则发现高的食糜黏度能够降低胃的排空速度,因此会引起饱感而减少采食量。果胶的天然黏性作用能够阻碍酶和胆汁盐反应,果胶与食糜混合后能扰乱食糜在小肠中的扩散,进而降低了营养物质的消化率[18]。Lagreca等[19]报道猪在不同时期添加果胶会产生不同的效果,饲粮中添加2%的果胶能显著降低早期日增重和饲料转化率,但是在育肥期添加4%果胶却显著提高日增重。Langhout等[20]在肉鸡玉米基础饲粮中添加3 mg/kg果胶,结果显著降低体增重、饲料转化率、空肠末端的能量和淀粉的利用率,其研究还发现添加果胶后增加了小肠食糜黏度,降低了小肠食糜的pH。Lin等[21]研究结果显示,添加果胶组中食糜黏度显著高于添加其他非淀粉多糖酶组,但是日采食量、日增重、饲料转化效率却显著低于添加其他非淀粉多糖酶组,添加果胶组胰腺组织中淀粉酶、脂肪酶、胰蛋白酶、糜蛋白酶活性显著低于添加其他非淀粉多糖酶组。
果胶酶的主要作用是补充内源酶的不足,刺激内源酶分泌,破坏植物细胞壁,促进营养物质消化吸收,消除抗营养因子,提高饲料营养价值[3]。Cowan等[25]研究了在14日龄肉鸡饲粮中添加单一果胶酶后,能量利用率和氨基酸消化率均有所提高,并且当饲喂低水平赖氨酸和蛋基酸饲粮时,添加果胶酶能够补偿氨基酸不足带来的影响。姜晓霞等[26]研究发现,饲粮中添加果胶酶能降解植物细胞壁的果胶,引起细胞壁裂解,有利于饲粮中粗纤维、酸性洗涤纤维和中性洗涤纤维的吸收和利用。许毅[27]研究发现,在玉米-豆粕型饲粮中添加果胶酶能显著提高肉鸡前期生产性能,改善全期生产性能,提高饲粮干物质、粗蛋白质和粗纤维消化率,果胶酶水平对十二指肠淀粉酶、总蛋白水解酶和空肠总蛋白水解酶活性的影响随果胶酶水平的提高有先升高后降低的趋势,但果胶酶水平对空肠淀粉酶和回肠食糜的黏度无显著影响。王春林等[28]研究发现,在玉米-豆粕饲粮中添加果胶酶能显著提高21日龄肉鸡的平均日增重和平均日采食量,并且提高饲料转化率,但在后期(21日龄后)添加果胶酶效果并不明显。
本试验中,添加果胶酶后肉鸡对2种豆粕、3种棉籽粕的代谢能和氨基酸消化率均得到不同程度的提高,菜籽粕的代谢能有所提高但不显著,这与上述报道中果胶酶能提高饲料养分利用率一致,其可能原因在于果胶酶破坏植物细胞壁后,释放了其中的一些可以部分利用的纤维素、半纤维素等多糖类物质,减少了排泄的粪尿能,因此提高了饲料代谢能。另外,果胶酶能够补充内源酶的不足[27],降低果胶带来的高食糜黏度[29],从而促进了酶对蛋白质和其他营养物质的消化,提高了氨基酸的消化率[30, 31]。虽然菜籽粕中的果胶含量高于豆粕和棉籽粕,但是本试验中添加果胶酶后改善了豆粕和棉籽粕的代谢能和氨基酸消化率,而菜籽粕却没有变化,可能原因在于果胶酶的作用不仅是分解果胶为还原糖,它还能改善肠道功能,提高内源酶活性[32],因此能够改善2种豆粕和3种棉籽粕的营养物质利用率,但是菜籽粕的纤维和抗营养因子大大高于豆粕和棉籽粕,它的阻碍效应可能让果胶酶的作用没能显现出来。Meng等[33]研究发现,添加复合酶制剂(包含果胶酶)后玉米饲粮、豆粕饲粮的总肠道非淀粉多糖消化率、蛋白质回肠消化率和AMEn显著提高,但是菜籽粕饲粮和豌豆饲粮没有显著作用,认为非淀粉多糖对营养物质的包裹作用可能不是影响饲粮营养物质利用率的唯一因素,因此果胶酶对菜籽粕的作用没有表现出来。Kocher等[34]在菜籽粕饲粮和向日葵粕饲粮中添加复合酶(葡聚糖酶、木聚糖酶、半纤维素酶和果胶酶)后,生长性能和AME无变化,但是空肠中总非淀粉多糖显著减少,其中菜籽粕型饲粮的AMEn显著降低,但原因不清楚。本试验结果与这2篇报道相符,推测这种结果也可能是由于果胶酶对菜籽粕的氨基酸的改善作用没有其抗营养因子的负面效应强而导致的。
本研究结果表明,同种原料经不同加工后形成的不同产品其氨基酸消化率和代谢能存在差异(优质豆粕优于普通豆粕,棉籽蛋白优于脱酚棉籽粕和普通棉籽粕),在饲粮中添加果胶酶能不同程度地改善肉鸡对饼粕类饲料原料的氨基酸消化率和代谢能。
| [1] | 刘文, 董赛丽, 梁金亚.果胶的性质、功能及其应用[J]. 三门峡职业技术学院学报, 2008, 7(2):118-121. ( 2) 2)
|
| [2] | 陈喜斌.饲料学[M]. 北京:科学出版社, 2003:12-13. (1)
|
| [3] | 周小乔, 王宝维.饲用果胶酶研究的进展[J]. 饲料研究, 2010(8):14-16. (3)
|
| [4] | SHORT F J, GORTON P, WISEMAN J, et al.Determination of titanium dioxide added as an inert marker in chicken digestibility studies[J]. Animal Feed Science and Technology, 1996, 59(4):215-221. (1)
|
| [5] | 邓雪娟, 刘国华, 蔡辉益, 等.分光光度计法测定家禽饲料和食糜中二氧化钛[J]. 饲料工业, 2008, 29(2):57-58. (1)
|
| [6] | 王永伟, 刘国华.家禽饲料有效能值的测定及影响因素[J]. 饲料工业, 2009, 30(11):41-44. (1)
|
| [7] | 张春雷, 侯水生, 刘福柱, 等.影响家禽饲料代谢能的因素[J]. 饲料工业, 2003, 24(8):11-13. (1)
|
| [8] | MATEOS G G, SELL J L.Influence of fat and carbohydrate source on rate of food passage of semipurified diets for laying hens[J]. Poultry Science, 1981, 60(9):2114-2119. (1)
|
| [9] | KETELS E, DE GROOTE G.Effect of ratio of unsaturated to saturated fatty acids of the dietary lipid fraction on utilization and metabolizable energy of added fats in young chicks[J]. Poultry Science, 1989, 68(11):1506-1512. (1)
|
| [10] | VILLAMIDE M J, SAN JUAN L D.Effect of chemical composition of sunflower seed meal on its true metabolizable energy and amino acid digestibility[J]. Poultry Science, 1998, 77(12):1884-1892. (1)
|
| [11] | 郑萍, 李波, 陈代文, 等.8种不同来源菜籽饼粕的生长猪氨基酸回肠消化率评定[J]. 中国畜牧杂志, 2012, 48(9):34-40. (1)
|
| [12] | 席鹏彬, 马永喜, 李德发, 等.中国菜籽饼粕化学组成特点及其影响因素的研究[J]. 中国畜牧杂志, 2004, 40(10):12-15. (1)
|
| [13] | 艾晓杰.家禽日粮抗营养因子的危害及对策[J]. 中国家禽, 2004, 26(1):8-11. (1)
|
| [14] | 龙玲, 吴达.饲料抗营养因子概述[J]. 饲料博览, 2001(7):12-16. (1)
|
| [15] | 王冲, 娄玉杰.常见饲料中抗营养因子及对动物的影响[J]. 家畜生态, 2000, 21(4):39-43. (2)
|
| [16] | 常磊.棉粕固态发酵及游离棉酚降解的研究[D]. 硕士学位论文.广州:暨南大学, 2012. (2)
|
| [17] | 荆丽珍, 王宝维, 魏笑笑, 等.绿色饲料添加剂——果胶酶的研究[J]. 饲料工业, 2007, 28(14):6-8. (1)
|
| [18] | 薛长湖, 张永勤, 李兆杰, 等.果胶及果胶酶研究进展[J]. 食品与生物技术学报, 2005, 24(6):94-99. (1)
|
| [19] | SCHNEEMAN B O.Soluble vs insoluble fiber:different physiological responses[J]. Food Technology, 1987, 41(2):81-82. (1)
|
| [20] | ROBERFROID M.Dietary fiber, inulin, and oligofructose:a review comparing their physiological effects[J]. Critical Reviews in Food Science & Nutrition, 1993, 33(2):103-148. (1)
|
| [21] | CHOCT M, ANNISON G.Anti-nutritive effect of wheat pentosans in broiler chickens:roles of viscosity and gut microflora[J]. British Poultry Science, 1992, 33(4):821-834. (1)
|
| [22] | LAGRECA L, MAROTTA E.Nutritional effect of pectin in swine during growth and before slaughter[J]. Arch Latinoam Nutrion, 1985, 35(1):172-179. (0)
|
| [23] | LANGHOUT D J, SCHUTTE J B, DE JONG J, et al.Effect of viscosity on digestion of nutrients in conventional and germ-free chicks[J]. British Journal of Nutrition, 2000, 83(5):533-540. (0)
|
| [24] | LIN P H, SHIH B L, HSU J C.Effects of different sources of dietary non-starch polysaccharides on the growth performance, development of digestive tract and activities of pancreatic enzymes in goslings[J]. British Poultry Science, 2010, 51(2):270-277. (0)
|
| [25] | COWAN W D, PETTERSSONI D R, RASMUSSEN P B.The influence of multi-component pectinase enzymes on energy and amino acid availability in vegetable proteins[C]//Proceedings of the Australian Poultry Science Symposium. Sydney:University of Sydney, 1999, 11:85-88. (1)
|
| [26] | 姜晓霞, 王宝维, 孙鹏, 等.鹅源草酸青霉产果胶酶对肉鸡生长发育影响研究[C]//中国家禽科学研究进展——第十四次全国家禽科学学术讨论会论文集.北京:中国畜牧兽医学会, 2009:737-743. (1)
|
| [27] | 许毅.果胶酶对肉鸡玉米豆粕型日粮养分利用率的影响[D]. 硕士学位论文.南京:南京农业大学, 2004. (2)
|
| [28] | 王春林, 陆文清, 李德发, 等.果胶酶对肉仔鸡生长性能及屠体性状的影响[J]. 中国畜牧杂志, 2004, 40(2):19-21. (1)
|
| [29] | 方华平, 程茂基.果胶酶在食品与饲料工业中的应用[J]. 饲料博览, 2006(5):34-35. (1)
|
| [30] | CHOCT M, HUGHES R J, WANG J, et al.Increased small intestinal fermentation is partly responsible for the anti-nutritive activity of non-starch polysaccharides in chickens[J]. British Poultry Science, 1996, 37(3):609-621. (1)
|
| [31] | MALATHI V, DEVEGOWDA G.In vitro evaluation of nonstarch polysaccharide digestibility of feed ingredients by enzymes[J]. Poultry Science, 2001, 80(3):302-305. (1)
|
| [32] | 王宝维, 姜晓霞, 孙鹏, 等.鹅源草酸青霉产果胶酶对肉鸡消化生理影响的研究[J]. 动物营养学报, 2010, 22(2):358-364. (1)
|
| [33] | MENG X, SLOMINSKI B A.Nutritive values of corn, soybean meal, canola meal, and peas for broiler chickens as affected by a multicarbohydrase preparation of cell wall degrading enzymes[J]. Poultry Science, 2005, 84(8):1242-1251. (1)
|
| [34] | KOCHER A, CHOCT M, PORTER M D, et al.The effects of enzyme addition to broiler diets containing high concentrations of canola or sunflower meal[J]. Poultry Science, 2000, 79(12):1767-1774. (1)
|