




后备牛生长发育关系到成年后奶牛的体型大小、泌乳及繁殖性能等的发挥,而中国后备奶牛的生长发育并未受到足够的重视,主要原因是缺乏统一的饲养管理标准、饲粮组成及营养水平差异较大、饲喂模式各不相同等。合理培育后备牛,不仅可延长奶牛的使用年限,提高饲料利用率和产奶量,增加经济效益,还有利于奶牛业的长期发展,为建立高产、稳产的优质奶牛群奠定基础。犊牛阶段适宜的平均日增重(ADG)、良好的饲料转化率以及健康的瘤胃和乳腺是后备牛培育的重点[1, 2]。有研究表明,奶牛在1~6月龄体重和体高相对增长速率最快,相对成本利用率最高[3]。故应充分利用犊牛阶段的生长优势,降低培育成本。提高饲粮营养水平可提高ADG,但过高的ADG会使初情期过早,减少生长母牛乳腺早期发育所用的时间,导致乳腺发育不够充分[4]。改变饲粮营养水平会引起血液内与营养代谢有关的代谢物质、酶活性、激素及生长因子等的变化,而这些变化又可作为代谢信号对机体生长发育(如体重、体尺和乳腺等)产生影响。通过测定生长发育指标与血清指标随饲粮能量水平的变化,研究不同能量水平的饲粮对后备牛生长发育的影响,对我国犊牛饲养实践中饲粮的配制具有重要指导意义。因此,本研究设定4种不同能量水平的饲粮,研究其对断奶犊牛生长发育和血清指标的影响,并以血清指标来指示犊牛健康、生长发育的优劣,尤其是以血清激素及生长因子含量来指示能量水平对乳腺发育潜在的可能影响,进而探讨断奶犊牛阶段饲粮适宜的能量水平,为犊牛培育提供参考。 1 材料和方法 1.1 试验设计与饲粮
试验采用单因素设计,选用32头完全断奶、健康的中国荷斯坦断奶母犊牛,按年龄、体重相近原则随机分成4组,每组8个重复,每个重复1头。正式试验开始时犊牛为(98.16±1.41)日龄,初始体重(91.80±1.82) kg。参照《中国奶牛饲养标准》(2004)[5]中125 kg生长母牛ADG为800 g营养需要量配制可消化粗蛋白质平均含量为9.30%,产奶净能分别为6.24(A组)、7.04(B组)、7.53(C组)和7.85 MJ/kg(D组)的4种试验饲粮(干物质基础)。试验饲粮组成及营养水平见表1。
| 表1 试验饲粮组成及营养水平(干物质基础) Table 1 Composition and nutrient levels of experimental diets (DM basis) % |
犊牛按组分圈饲养,保证牛舍卫生。每日饲喂2次(09:00和15:00),精料和玉米青贮按比例混合饲喂,苜蓿干草单独饲喂。预试期10 d,预试期各组试验牛均饲喂B组饲粮(产奶净能为7.04 MJ/kg,可消化粗蛋白质为9.28%)。正试期82 d,各组试验牛日饲喂量相同,约为体重的3%,每2周根据采食情况和体重变化进行调整。自由饮水。
1.3 测定指标与方法 1.3.1 生长发育指标在98、120、150和180日龄晨饲前测定每头犊牛的体重、体尺、前乳头长度和后乳头长度,并进行体况评分,根据各生长阶段始末体重计算ADG。详细记录每天每组犊牛的饲喂量和剩料量,每2周采集饲料和剩料样品测定营养成分,计算各组干物质采食量和饲料转化率。乳头长度和体况评分测定参照Lammers等[6]和Wildman等[7]的方法。
1.3.2 血清指标犊牛称重前每组随机选取4头采集血液,使用真空管颈静脉穿刺采血约20 mL,3 000 r/min离心20 min,分离血清于1.5 mL离心管,-20 ℃保存。血清样品采用全自动生化仪(日立7160,日本)和酶联免疫吸附测定法(ELISA)试剂盒测定相关指标。测定指标包括血清生化指标:葡萄糖(GLU)、总蛋白(TP)、白蛋白(ALB)、球蛋白(GLB)、尿素氮(UN)、胆固醇(CHO)、甘油三酯(TG)、高密度脂蛋白(HDL)、低密度脂蛋白(LDL)和游离脂肪酸(FFA)含量及总抗氧化能力(T-AOC);血清激素及生长因子:生长激素(GH)、雌二醇(E2)、胰岛素(INS)、胰高血糖素(GC)、胰岛素样生长因子-Ⅰ(IGF-Ⅰ)和瘦素含量。
1.4 统计分析采用SAS 9.1.3统计软件中的单因素方差分析(one-way ANOVA)和关于重复测量数据的MIXED模型进行分析,并采用LSD法进行双重比较,统计模型如下:
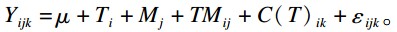
式中:μ为平均数;T为处理(i=1、2、3、4),固定效应;M为出生后日龄(j=1、2、3、4),固定效应;C为犊牛(k=1,2···32),随机效应;ε为残差。
统计分析以P<0.05为差异显著,P<0.01为差异极显著,P在0.05~0.10之间为有提高或降低的趋势。
2 结果与分析 2.1 犊牛生长发育 2.1.1 体重和ADG由表2可知,在98日龄,犊牛起始体重各组间无显著差异(P>0.05),平均值为91.80 kg;在120和150日龄,犊牛体重各组间差异不显著(P>0.05);在180日龄,犊牛体重随饲粮中产奶净能的增加呈升高的趋势(P<0.10),D组犊牛体重分别比A、B和C组提高了11.65%、5.32%和3.69%。在试验全期(98~180日龄),犊牛ADG各组间差异不显著(P>0.05),但D组ADG达到0.84 kg/d,与A、B和C 3组相比,分别提高了31.25%、12.00%和7.69%;在98~120日龄和121~150日龄,犊牛ADG各组间差异不显著(P>0.05),A组略低于其他各组;在151~180日龄,D组犊牛ADG极显著高于A组(P<0.01),显著高于C组(P<0.05),且有高于B组的趋势(P<0.10)。
| 表2 不同能量水平的饲粮对犊牛体重和ADG的影响 Table 2 Effects of diets with different energy levels on body weight and ADG of calves
|
随日龄增加,犊牛的体重和ADG均出现极显著的变化(P<0.01),试验处理与日龄间交互作用不显著(P>0.05)。
2.1.2 干物质采食量和饲料转化率由表3可知,犊牛精料、苜蓿干草、玉米青贮和总干物质采食量各组间均无显著差异(P>0.05),平均总干物质采食量为3.64 kg,B组总干物质采食量数值上均略高于其他各组;C和D组犊牛的饲料转化率得到显著改善(P<0.05),饲料转化率数值分别比A组降低了18.72%和29.29%。
| 表3 不同能量水平的饲粮对犊牛干物质采食量和饲料转化率的影响 Table 3 Effects of diets with different energy levels on DMI and feed conversion ratio of calves |
由表4可知,在试验全期及各阶段,犊牛体高各组间均无显著差异(P>0.05)。在试验全期、98、120和150日龄,犊牛体斜长各组间均无显著差异(P>0.05);不同能量水平的饲粮对180日龄犊牛体斜长影响显著(P<0.05)。不同日龄犊牛的胸围、腹围和管围组间差异均不显著(P>0.05)。在试验全期及各阶段,犊牛腰角宽组间差异均不显著(P>0.05),但在180日龄,犊牛腰角宽随饲粮中产奶净能的增加有升高的趋势(P<0.10)。
| 表4 不同能量水平的饲粮对犊牛体尺的影响 Table 4 Effects of diets with different energy levels on body size of calves cm |
试验全期犊牛各体尺指标均随日龄的增加极显著升高(P<0.01),试验处理与日龄间无显著交互作用(P>0.05)。
2.1.4 体况评分由表5可知,在180日龄,犊牛体况评分变化范围在2.41~2.75之间,D组犊牛体况评分极显著高于A组(P<0.01),且D组体况评分在整个试验期提高了0.63,增长值高于其他3组。试验全期犊牛体况评分随日龄增加极显著升高(P<0.01),试验处理与日龄间交互作用不显著(P>0.05)。
| 表5 不同能量水平的饲粮对犊牛体况评分的影响 Table 5 Effects of diets with different energy levels on body condition score of calves |
由表6可知,在试验全期,犊牛4个乳头长度受饲粮能量水平影响均不显著(P>0.05),前后乳头长度在1.87~2.11 cm和1.67~1.86 cm之间变化,在试验全期,犊牛前乳头平均长度要比后乳头平均长度长0.20 cm。试验全期犊牛乳头长度随日龄增加极显著提高(P<0.01),试验处理与日龄间的交互作用不显著(P>0.05)。
| 表6 不同能量水平的饲粮对犊牛乳头长度的影响 Table 6 Effects of diets with different energy levels on nipple length of calves cm |
2.2 犊牛血清指标 2.2.1 血清生化指标
由表7可知,在试验全期,犊牛血清GLU含量各组间均无显著差异(P>0.05),维持在5.77~6.17 mmol/L之间变化。在150和180日龄,犊牛血清GLU含量随饲粮能量水平的增加呈升高趋势(P<0.10)。在120日龄,犊牛血清TP、ALB和GLB含量均出现显著变化(P<0.05),且在150日龄,B组犊牛血清ALB含量显著低于A、C和D 3组(P<0.05),其他日龄组间无显著差异(P>0.05)。在试验全期、120和150日龄,犊牛血清UN含量均有随饲粮能量水平提高而降低的趋势(P<0.10)。犊牛血清T-AOC在试验各阶段组间均无显著差异(P>0.05)。
| 表7 不同能量水平的饲粮对犊牛血清生化指标的影响 Table 7 Effects of diets with different energy level on serum biochemical indexes of calves |
在试验全期,C和D组犊牛血清CHO含量显著高于A和B组(P<0.05),并随饲粮能量水平的提高逐渐升高;在98日龄,犊牛血清CHO含量各组间无显著差异(P>0.05),在150和180日龄极显著升高(P<0.01)。犊牛血清TG含量在试验各阶段组间差异均不显著(P>0.05)。在试验全期,犊牛血清HDL和LDL含量组间差异均不显著(P>0.05),但分别在150和180日龄、120和150日龄出现显著变化(P<0.05),且在180日龄C和D组血清HDL含量显著高于A组(P<0.05)。在98日龄,犊牛血清FFA含量各组间差异显著(P<0.05),并在180日龄呈降低趋势(P<0.10)。
在试验全期,犊牛血清GLU、CHO、TG、HDL、LDL和UN含量随日龄的增加极显著升高(P<0.01),ALB含量随日龄的增加显著升高(P<0.05),FFA、TP、GLB含量和T-AOC受日龄影响不显著(P>0.05)。犊牛血清ALB含量受试验处理和日龄交互作用影响显著(P<0.05),其他血清指标含量的试验处理与日龄交互作用不显著(P>0.05)。 2.2.2 血清激素及生长因子
由表8可知,犊牛血清GH含量整体在数值上逐渐减低,并在120日龄组间达到显著水平(P<0.05)。犊牛血清E2含量在120和180日龄分别出现极显著(P<0.01)和显著(P<0.05)变化,并随饲粮中产奶净能的增加呈先升高后降低的变化,且B组含量最高。在整个试验期,饲粮中产奶净能的增加对犊牛血清INS含量有显著影响,D组INS含量显著低于A、B和C 3组(P<0.05),特别是在120日龄,D组极显著或显著低于A、B和C组(P<0.01、P<0.01和P<0.05)。在试验各阶段,犊牛血清GC含量各组间差异均不显著(P>0.05)。在120日龄,D组犊牛血清IGF-Ⅰ含量低于A、B和C组(P>0.05、P<0.01和P<0.05),其他阶段变化不显著(P>0.05)。在180日龄,B组犊牛血清瘦素含量显著高于A、C和D组(P<0.05),其他阶段各组间无显著差异(P>0.05)。
| 表8 不同能量水平的饲粮对犊牛血清激素及生长因子的影响 Table 8 Effects of diets with different energy levels on hormones and growth factors in serum of calves |
试验全期犊牛血清GC和IGF-Ⅰ含量随日龄的增加显著升高(P<0.05),INS含量随日龄的增加极显著升高(P<0.01),而GH、E2和瘦素含量不受日龄因素影响(P>0.05)。上述指标试验处理和日龄的交互作用均不显著(P>0.05)。 3 讨 论 3.1 不同能量水平的饲粮对犊牛生长发育的影响
1~6月龄中国荷斯坦牛生长发育强度依次为体重、体长、胸围和体高[8]。采食量高低影响动物的生长发育[1],能量摄入水平随干物质采食量的升高而升高,影响犊牛ADG。本试验结果表明,在试验全期,犊牛总干物质采食量各组间差异不显著,保持在3.43~3.77 kg之间。在试验全期,C和D组犊牛饲料转化率得到显著改善,较A组分别降低了1.08和1.69,提高了饲粮利用效率。Davis等[9]研究表明,3~6月龄犊牛采食低(代谢能为9.37 MJ/kg)和高(代谢能为11.80 MJ/kg)能量水平饲粮的干物质采食量占体重的比值(DMI/BW)分别为2.8%和3.3%,ADG分别为0.64和1.09 kg/d。本研究中各组犊牛产奶净能水平依次为6.24、7.04、7.53和7.85 MJ/kg,其TDMI/BW分别为3.2%、3.2%、3.0%和2.8%,ADG分别为0.64、0.75、0.78和0.84 kg/d,ADG随TDMI/BW降低而升高,原因可能是饲粮能量水平不同而采食量相近引起的。
表2结果显示,试验期犊牛初始体重和末体重各组间无显著差异,但在180日龄,犊牛体重出现升高的趋势,且在151~180日龄,D组犊牛ADG极显著高于A组,达到1.24 kg/d,生长速度过快。Kertz等[3]研究表明,断奶后的后备奶牛ADG保持在0.82~0.93 kg/d范围内体重可呈线性增长,提高生长效率,降低培育成本。本试验在121~150日龄,犊牛ADG在0.76~0.86 kg/d之间,基本达到上述推荐范围。在151~180日龄,犊牛ADG受不同能量水平饲粮影响出现极显著变化;在180日龄,犊牛体斜长显著增加,体重及腰角宽呈升高的趋势,这与该生长阶段犊牛生长发育强度相一致。Kertz等[3]建议体高在6月龄时达到104 cm,与本试验180日龄(6月龄)结果相一致,表明试验设定的饲粮能量水平能满足犊牛生长需要。
本试验犊牛在180日龄以前,体况评分各组间无显著差异。在180日龄,D组犊牛体况评分显著高于A组,同时,C组犊牛的体况评分全期增长0.30。D组体况评分的提高与本阶段高ADG、体尺增长和体内脂肪的沉积等密切相关。Shamay等[10]研究结果显示,犊牛体重在150~200 kg,体况评分范围在2.00~2.50之间,本试验180日龄犊牛体况评分在2.41~2.75之间,稍高于前者,表明试验饲粮能量水平的提高增加了犊牛的体况评分。
3.2 不同能量水平的饲粮对犊牛血清生化指标的 影响能量水平显著影响犊牛ADG、血液生理生化指标和血清电解质离子含量[11]。断奶后犊牛不能或很少直接消化饲粮中碳水化合物获得机体所需GLU,而是通过糖异生途径主要由丙酸合成。研究表明,3~12月龄后备牛血清GLU含量随饲粮能量水平升高而升高[12],而本试验结果显示,4组血清GLU含量在5.77~6.17 mmol/L之间,各组间差异不显著,可能因为组间能量水平不足以引起GLU含量的显著变化。在120日龄,犊牛血清GLU含量在整个试验期较低,原因可能与冬季饲养环境温度较低,犊牛饮用冰水引起微生物数量减少,发酵生成挥发性脂肪酸含量降低,从而造成丙酸糖异生为糖的含量下降有关。
本研究TG含量组间无显著差异,这与张蓉等[13]的试验结果一致,表明提高产奶净能水平对脂肪沉积影响不显著。但本研究中犊牛血清TG含量随日龄增加极显著增加,试验全期为0.74 mmol/L,远大于其1~2月龄0.13 mmol/L,表明脂肪沉积与年龄的相关性较大,犊牛阶段能量主要用于快速生长而非脂肪的沉积。本试验能量水平极显著提高试验全期犊牛血清CHO含量,显著提高HDL和LDL含量,且CHO、HDL和LDL含量升高趋势相同,而FFA含量随能量水平增加呈降低的趋势。由于血液中CHO主要由LDL携带运输,HDL有助于清除细胞内CHO,三者含量之间存在关联,所以,血清含量变化趋势相同。3~12月龄后备牛血清FFA含量随能量水平升高而降低[12],与本试验研究结果相近。综上,能量水平改变了血清内与脂肪代谢相关的代谢产物,但对机体健康没有造成不良影响。
血清UN作为犊牛体内氮代谢一项重要衡量指标,可反映饲粮蛋白质和氨基酸水平。采食高蛋白质(26%)代乳粉提高早期断奶犊牛的血清UN含量[14],而增加代乳粉脂肪水平(113 g/d)则会降低犊牛血清UN含量[15]。本试验犊牛血清UN含量随能量水平的升高有降低的趋势,这可能因为添加脂肪提高能量水平影响了机体的氮代谢。
对犊牛全期血清TP含量进行分析,所有组犊牛血清TP含量基本稳定在63.00 g/L,表明犊牛机体摄入蛋白质的量变化不大。本试验在试验全期,犊牛血清ALB和GLB含量分别在27.79~28.07 g/L和34.21~35.90 g/L之间,属正常范围。T-AOC代表了清除体内活性氧的抗氧化物水平,本试验全期犊牛血清T-AOC不受饲粮能量水平的影响,稳定在8.50~9.05 U/mL之间。在120日龄,犊牛血清TP、ALB和GLB含量均出现显著变化,可能由于环境因素(此试验期是北京冬天最冷的时候)造成犊牛产生冷应激,导致机体营养物质代谢紊乱及免疫力降低。在150日龄,B组犊牛血清ALB含量显著低于A、C和D 3组,CHO、HDL和LDL含量在150日龄均出现了较大的变化,可能是饲粮脂肪酸代谢引起上述指标的变化。同时,日龄的增加及试验处理与日龄的交互作用均显著影响血清ALB含量。以上结果表明,除去环境因素的影响,试验设定的不同能量水平的饲粮均能保证犊牛的健康,对犊牛免疫力影响不显著。
3.3 不同能量水平的饲粮对犊牛血清激素及生长因子的影响机体内激素及生长因子调控动物机体的生长发育。INS通过促进GLU转化合成脂肪,降低GLU含量并促进蛋白质的合成;INS和GC共同调节机体GLU代谢、蛋白质合成代谢,维持GLU含量的稳定[16]。研究指出,妊娠母羊血清GLU与INS含量呈显著正相关[17]。本研究结果显示,在试验全期,D组犊牛血清INS含量显著低于A、B和C 3组,而GC含量各组间差异不显著,且日龄的影响要大于饲粮处理。在120日龄,A和B组血清INS含量极显著高于D组,这可能与血清GLU含量在120日龄低于其他日龄有关,犊牛需提高分解代谢以维持机体需要。
采食高营养水平饲粮可显著降低血清GH含量[18],荷斯坦奶牛的体重及ADG与血清GH含量显著相关[16, 19, 20]。注射外源牛生长激素能显著增加青年母牛ADG[21, 22],改善饲料转化率,降低机体脂肪沉积率[23]。IGF-Ⅰ在生长发育过程中同样具有重要作用,其合成与分泌受血液GH含量的调节,形成一个GH-IGF轴,共同调控机体内大多数组织的生长发育[23]。Chelikani等[12]研究3~12月龄后备牛血清IGF-Ⅰ含量随饲粮能量水平升高而升高,初情期前母猪受到能量限饲的影响,血清IGF-Ⅰ含量降低[24]。本试验犊牛血清GH含量逐渐降低,并在120日龄时达到显著水平;B组血清IGF-Ⅰ含量在120日龄极显著高于D组,A和C组均显著高于D组,与GH含量变化趋势相同,这与98~120日龄低ADG相对应。血清GH含量随ADG的升高而降低,并随饲粮能量水平的变化逐渐降低。
瘦素主要由脂肪组织产生,对哺乳动物机体能量代谢调节极为关键,循环血中瘦素与机体脂肪含量成反比[25]。Brown等[26]研究表明,饲粮营养可调控血浆瘦素含量,4~6周龄犊牛血浆瘦素含量随营养水平升高而升高。Chelikani等[12]研究也表明,在34周龄,生长母牛血浆瘦素含量受饲粮处理影响出现显著升高(低、高营养组分别为2.81和3.86 μg/L)。而Block等[27]的研究结果显示,饲粮脂肪来源(棕榈酸钙盐和共轭亚油酸)对3~5月龄犊牛ADG和血浆瘦素含量没有显著影响,并指出奶牛1岁前血浆瘦素含量会一直维持在2.3 μg/L。而Chelikani等[12]研究结果显示,血浆瘦素含量随日龄增加显著升高,而初情期母牛瘦素含量在不同ADG组(1.10、0.80和0.50 kg/d)间差异不显著。本研究结果显示,试验全期犊牛血清瘦素含量受日龄影响不显著;在试验全期,其平均含量为9.17 μg/L,高于上述研究结果,可能与饲粮添加高脂肪水平有关;在180日龄,B组犊牛血清瘦素含量显著高于A、C和D组,B、C和D组血清瘦素含量随饲粮能量水平变化(过瘤胃脂肪添加量升高)显著降低,与该阶段采食量的提高、机体脂肪沉积量的增加有关;A组含量低于其他各组,可能是因为低能量水平饲粮营养主要供机体维持与生长,脂肪沉积量很少导致。
3.4 不同能量水平的饲粮对犊牛乳腺发育的影响奶牛头胎产奶量与犊牛阶段体高、体长[28]、乳房外观尺寸[6, 11, 28]和初产体重呈显著正相关[29],要达到理想初产体重,需要后备牛阶段ADG达到一定的水平,但过快的ADG容易对乳腺发育产生不利影响,进而降低产奶量[30]。有学者指出,产前ADG超过700 g/d就会对乳腺发育和成年后的产奶量产生影响[6 ,9,31],但也有研究表明提高ADG(725~950 g/d)不会影响产奶量[32]。本试验结果表明,前后乳头长度随日龄增加而显著增加,各组间并无显著差异,可能因为4组饲粮差异不显著或是受到年龄、ADG、试验期和饲粮其他因素等的限制,这与张卫兵等[33]不同蛋白质能量比的饲粮对3~5月龄犊牛乳头长度的影响结果一致。
乳腺发育受某些饲粮营养水平、饲粮诱导的代谢和激素等变化的影响较大[34],饲粮能量可通过激素及生长因子调节这些代谢过程调控乳腺发育。本试验犊牛全期血清GH含量在数值上逐渐降低,表明饲粮能量水平的提高会降低GH含量,进而对乳腺发育产生影响。研究指出,乳腺发育与GH含量正相关[18],9月龄前生长母牛采食过高营养影响乳腺发育,其原因可能是高营养水平导致血清GH含量降低[4]。本研究中,犊牛血清E2含量在试验全期不受饲粮因素影响,与张卫兵等[33]的3~5月龄不同蛋能比试验结果一致,可能因为该生长阶段处于快速生长发育期,因未达到初情期体内E2含量较低,对乳腺发育影响较小。本试验全期犊牛血清IGF-Ⅰ含量D组最低,瘦素含量B组最高。IGF-Ⅰ可促进细胞分化,瘦素可防止乳腺脂肪过多沉积,在一定程度上表明高能量水平影响乳腺的正常发育。前人研究一致认为IGF-Ⅰ和瘦素等在乳腺发育过程中发挥了重要的作用[27, 35],但其对乳腺发育的调节机制尚待研究[9]。
4 结 论在本试验条件下,提高饲粮能量水平:
① 可显著改善犊牛饲料转化率;在180日龄,饲喂产奶净能为7.85 MJ/kg的饲粮可显著提高犊牛ADG、体斜长和体况评分。
② 可提高犊牛血清CHO含量,影响LDL和UN含量变化趋势。
③ 对犊牛血清E2、瘦素和INS含量产生影响。
综合本试验各指标,3~6月龄犊牛饲喂产奶净能为7.53 MJ/kg的饲粮为宜。
| [1] | SEJRSEN K,HUBER J T,TUCKER H A,et al.Influence of nutrition on mammary development in pre-and post-pubertal heifers[J]. Journal of Dairy Science,1982,65(5):793-800. ( 2) 2)
|
| [2] | 张卫兵,刁其玉,赵广城.从营养和饲养角度加强后备牛的培育[J]. 乳业科学与技术,2008,31(1):24-27. (1)
|
| [3] | KERTZ A F,BARTON B A,REUTZEL L F.Relative efficiencies of wither height and body weight increase from birth until first calving in Holstein cattle[J]. Journal of Dairy Science,1998,81(5):1479-1482. (3)
|
| [4] | MEYER M J,CAPUCO A V,ROSS D A,et al.Developmental and nutritional regulation of the prepubertal bovine mammary gland:Ⅱ.Epithelial cell proliferation,parenchymal accretion rate,and allometric growth[J]. Journal of Dairy Science,2006,89(11):4298-4304. (2)
|
| [5] | 冯仰廉,方有生,莫放,等.中国奶牛饲养标准[M].北京:中国农业出版社,2004. (1)
|
| [6] | LAMMERS B P,HEINRICHS A J,KENSINGER R S.The effects of accelerated growth rates and estrogen implants in prepubertal Holstein heifers on estimates of mammary development and subsequent reproduction and milk production[J]. Journal of Dairy Science,1999,82(8):1753-1764. (3)
|
| [7] | WILDMAN E E,JONES G M,WAGNER P E,et al.A dairy cow body condition scoring system and its relationship to selected production characteristics[J]. Journal of Dairy Science,1982,65(3):495-501. (1)
|
| [8] | 李文,刘小林,文利侠,等.中国荷斯坦后备奶牛生长发育规律的研究[J]. 畜牧兽医杂志.2007,26(5):19-21,24. (1)
|
| [9] | DAVIS RINCKER L E,NIELSEN W,CHAPIN L T,et al.Effects of feeding prepubertal heifers a high-energy diet for three,six,or twelve weeks on feed intake,body growth,and fat deposition[J]. Journal of Dairy Science,2008,91(5):1913-1925. (3)
|
| [10] | SHAMAY A,WERNER D,MOALLEM U,et al.Effect of nursing management and skeletal size at weaning on puberty,skeletal growth rate,and milk production during first lactation of dairy heifers[J]. Journal of Dairy Science,2005,88(4):1460-1469. (1)
|
| [11] | JABBAR L,CHEEMA A M,JABBAR M A,et al.Effect of different dietary energy levels,season and age on hematological indices and serum electrolytes in growing buffalo heifers[J]. The Journal of Animal and Plant Sciences,2012,22:279-283. (2)
|
| [12] | CHELIKANI P K,AMBROSE D J,KEISLER D H,et al.Effects of dietary energy and protein density on plasma concentrations of leptin and metabolic hormones in dairy heifers[J]. Journal of Dairy Science,2009,92(4):1430-1441. (5)
|
| [13] | 张蓉,刁其玉,屠艳,等.能量水平对早期断奶犊牛消化代谢及血清指标的影响[J]. 中国农业科学,2009,42(3):1024-1029. (1)
|
| [14] | 李辉,刁其玉,张乃锋,等.不同蛋白水平对犊牛消化代谢及血清生化指标的影响[J]. 中国农业科学,2008,41(4):1219-1226. (1)
|
| [15] | JASTER E H,MCCOY G C,FERNANDO R L.Dietary fat in milk or milk replacers for dairy calves raised in hutches during the winter[J]. Journal of Dairy Science,1990,73(7):1843-1850. (1)
|
| [16] | 刘扬,昝林森,林立亚,等.营养条件对荷斯坦公犊生长发育及血液中激素水平的影响[J]. 中国畜牧杂志,2009,45(5):25-30. (2)
|
| [17] | 冷静,王刚,朱仁俊,等.不同补饲水平对妊娠各期努比亚母羊激素及血液生化指标的影响[J]. 中国畜牧兽医,2011,38(2):21-24. (1)
|
| [18] | SEJRSEN K,HUBER J T,TUCKER H A.Influence of amount fed on hormone concentrations and their relationship to mammary growth in heifers[J]. Journal of Dairy Science,1983,66(4):845-855. (2)
|
| [19] | ARMSTRONG D T,HANSEL W.The effect of age and plane of nutrition on growth hormone and thyrotropic hormone content of pituitary glands of Holstein heifers[J]. Journal of Animal Science,1956,15(3):640-649. (1)
|
| [20] | PURCHAS R W,MACMILLAN K L,HAFS H D.Pituitary and plasma growth hormone levels in bulls from birth to one year of age[J]. Journal of Animal Science,1970,31(2):358-363. (1)
|
| [21] | VESTERGAARD M,PURUP S,HENCKEL P,et al.Effects of growth hormone and ovariectomy on performance,serum hormones,insulin-like growth factor-binding proteins,and muscle fiber properties of prepubertal Friesian heifers[J]. Journal of Animal Science,1995,73(12):3574-3584. (1)
|
| [22] | RADCLIFF R P,VANDEHAAR M J,CHAPIN L T,et al.Effects of diet and injection of bovine somatotropin on prepubertal growth and first-lactation milk yields of Holstein cows[J]. Journal of Dairy Science,2000,83(1):23-29. (1)
|
| [23] | 张童非,崔英俊,李庆章.IGF-Ⅰ在奶牛乳腺的表达及其对乳腺上皮细胞泌乳功能的影响[J]. 中国畜牧兽医,2012,39(5):126-129. (2)
|
| [24] | 刘立文,张立永,高志花,等.能量对初情期前母猪类胰岛素样生长因子分泌的影响[J]. 黑龙江畜牧兽医,2008(4):23. (1)
|
| [25] | 顾卫琼,陈名道,唐金凤.中国人血清瘦素水平与肥胖度的关系[J].中华内分泌代谢杂志,1999,15(1):15. (1)
|
| [26] | BROWN E G,VANDEHAAR M J,DANIELS K M,et al.Effect of increasing energy and protein intake on body growth and carcass composition of heifer calves[J]. Journal of Dairy Science,2005,88(2):585-594. (1)
|
| [27] | BLOCK S S,SMITH J M,EHRHARDT R A,et al.Nutritional and developmental regulation of plasma leptin in dairy cattle[J]. Journal of Dairy Science,2003,86(10):3206-3214. (2)
|
| [28] | 刘正伦,唐宗南,蒋炯琳.犊牛乳腺生长发育和外貌性状与产乳量的相关研究[J]. 中国奶牛,1984(1):8-14. (2)
|
| [29] | LIN C Y,LEE A J,MCALLISTER A J,et al.Intercorrelations among milk production traits and body and udder measurements in Holstein heifers[J]. Journal of Dairy Science,1987,70(11):2385-2393. (1)
|
| [30] | SEJRSEN K,PURUP S,VESTERGAARD M,et al.High body weight gain and reduced bovine mammary growth:physiological basis and implications for milk yield potential[J]. Domestic Animal Endocrinology,2000,19(2):93-104. (1)
|
| [31] | VAN AMBURGH M E,GALTON D M, BAUMAN D E,et al.Effects of three prepubertal body growth rates on performance of Holstein heifers during first lactation[J]. Journal of Dairy Science,1998,81(2):527-538. (1)
|
| [32] | WALDO D R,CAPUCO A V,REXROAD C E,Jr.Milk production of Holstein heifers fed either alfalfa or corn silage diets at two rates of daily gain[J]. Journal of Dairy Science,1998,81(3):756-764. (1)
|
| [33] | 张卫兵,刁其玉,张乃锋,等.日粮蛋白能量比对3-5月龄后备奶牛乳腺发育及相关激素的影响[J]. 中国畜牧杂志,2011,47(7):46-49. (2)
|
| [34] | BERRYHILL G E,GLOVICZKI J M,TROTT J F,et al.Diet-induced metabolic change induces estrogen-independent allometric mammary growth[J]. Proceedings of the National Academy of Sciences of the United States of America,2012,109(40):16294-16299. (1)
|
| [35] | GARCIA M R,AMSTALDEN M,MORRISON C D,et al.Age at puberty,total fat and conjugated linoleic acid content of carcass,and circulating metabolic hormones in beef heifers fed a diet high in linoleic acid beginning at four months of age[J]. Journal of Animal Science,2003,81(1):261-268. (1)
|