




2. 江西省职业病防治研究院, 南昌 330000
2. Institute of Occupational Medicine of Jiangxi, Nanchang 330000, China
蜜蜂是与人类以及大自然关系密切的社会性昆虫,它不仅可以为人类提供营养丰富的蜂产品,而且通过授粉模式维持生态平衡,对保护和维持生物多样性起着重要作用[1]。蜂王是蜜蜂蜂群当中的主要繁殖个体,蜂王质量决定群势的强弱,高质量蜂王对感染寄生虫病有较高的抵抗能力,因此培育高质量、高抵抗力的蜂王对于养蜂生产尤为重要。在传统育王过程中,养蜂人员通常通过奖励饲喂糖水来提高蜂王的培育质量,仅依靠外界粉源及巢脾中储存的蜂粮提供蛋白质营养。但由于季节等因素,外界粉源缺乏、蜂群自身储备又不足时,会导致蜜蜂营养物质的缺乏,此时会对蜜蜂的繁殖及蜂王的培养产生较大的影响,有经验的养蜂人员在缺粉的情况下会适当添加一些花粉,但不同花粉的营养效价不一样,而且随着花粉价格的不断提升,许多养蜂人员会为了降低养蜂成本而选择黄豆粉等人工合成饲粮来代替花粉,而人工合成饲粮中蛋白质水平不同对蜂群的应用效果也不一样。近年来,蜜蜂蛋白质营养需要研究取得了阶段性成果,研究发现:蛋白质缺乏会影响蜜蜂的生长发育,导致蜂王产卵和工蜂育子能力降低,影响羽化工蜂的体重和体组织蛋白质含量、产浆量、卵的孵化率、幼虫化蛹率、机体抗氧化活性,使蜂群群势急剧下降,甚至导致整个蜂群消亡等[2-9]。目前关于饲粮蛋白质水平对育王质量的影响未见报道。基于此,我们开展了本项试验,通过探索饲粮粗蛋白质水平对中华蜜蜂(Apis cerana cerana)育王质量的影响,为优质蜂王培育提供一定的理论基础。
1 材料与方法 1.1 试验动物试验动物为江西农业大学蜜蜂研究所饲养的中华蜜蜂,于2015年8月至10月在江西农业大学蜜蜂研究所开展试验。
1.2 饲粮及主要试剂不同粗蛋白质水平的饲粮(由山东农业大学提供),油菜蜜,DNA聚合Taq酶和对应的缓冲液Buffer(北京康为世纪生物科技有限公司产品),Trizol总RNA提取试剂盒和RNA酶抑制剂(北京全式金生物技术有限公司产品),Oligo(dT)(5.97 nmol/OD,由美国英杰生命技术公司上海办事处合成),dNTP Mixture(2.5 mmole,TaKaRa),反转录酶M-mLV(200 U/μL,TaKaRa)。
1.3 主要仪器生化培养箱(GZ-250-GSI型,韶关市广智科技设备发展有限公司产品),核酸蛋白质测定仪 (NanoPhotometerTM P300,IMPLEN),普通离心机(飞鸽KA-1000型,上海安亭科学仪器厂公司),荧光定量PCR仪(iQTM2型,Bio-Rad公司),PCR仪(iQTM2型,Bio-Rad公司)。
1.4 试验方法 1.4.1 蜜蜂饲喂及育王选取群势相当的中华蜜蜂9群(每群5脾蜂),随机分成3组,每组3群,在每群蜂巢门口装置脱粉器,阻止花粉进入,使其处于缺粉状态。Ⅰ组、Ⅱ组、Ⅲ组蜂群分别饲喂粗蛋白质水平为15%、25%、35%的试验饲粮,试验饲粮组成及营养水平见表 1[7],每隔3 d饲喂1次,每次足量饲喂。定期对蜂群进行检查并饲喂糖水。饲喂45 d后,将提前准备好的空巢脾分别添加到每群蜂中,控制蜂王产卵8 h后,用隔王板将产卵的巢脾和蜂王分开。3 d后,按照标准的人工育王方法培育中华蜜蜂蜂王[10]。
|
|
表 1 试验饲粮组成及营养水平(风干基础) Table 1 Composition and nutrient levels of experimental diets (air-dry basis) |
在处女王出生的前1天,将王台按组别统一装入框式王笼中,移至恒温恒湿培养箱(温度34 ℃,相对湿度80%)中羽化出房,用分析天平称量刚出房蜂王的体重,同时测量其胸宽,记录试验数据。
1.4.3 蜂王卵巢卵黄蛋白原基因(vitellogenin,Vg)、转铁蛋白基因(transferrin,Trf)表达的测定 1.4.3.1 样品采集刚出房的蜂王测完初生重、胸宽后立即解剖,采集卵巢,分别装入1.5 mL RNase-free的EP管迅速放入液氮中,用于后续检测。
1.4.3.2 总RNA的提取与cDNA的合成每组取4只蜂王的卵巢,放入含液氮的研钵中进行研磨,研磨成粉末后转入装有1 mL Trizol的1.5 mL的EP管中。参考秦秋红[11]的方法对样本总RNA进行提取,用核酸蛋白质测定仪测定其OD260/280(1.9~2.1之间符合标准)和总RNA浓度,每个组测3次,取平均值。
用反转录试剂盒对总RNA进行反转录,反应体系为50 μL,包括:8 μL总RNA,10 μL Buffer,8 μL dNTP,1.5 μL反转录酶M-mLV,3 μL Oligo(dT),1 μL RNA酶Inhibitor,18.5 μL焦碳酸二乙酯(DEPC)水。反转录反应条件如下:体系混匀后,42 ℃反应60 min,75 ℃灭活5 min。反转录产物保存于-80 ℃。
1.4.3.3 荧光定量PCR引物的设计及荧光定量PCR参照从东方蜜蜂卵巢组织中克隆的基因mRNA序列(Vg:AJ517411;Trf:AY336529.1) 及GenBank数据库中中华蜜蜂的转录组序列[12],用Primer 5.0软件设计引物序列(表 2),以β-肌动蛋白(β-actin)作为内参基因。荧光定量PCR反应体系(10 μL)如下:cDNA 1 μL,SYBR Premix ExTaqTM Ⅱ 5 μL,上游和下游引物各0.4 μL,超纯灭菌水3.2 μL。体系混匀,放入CFX96定量仪中进行扩增。反应条件:95 ℃ 30s,95 ℃ 10 s,60 ℃ 1 min,40个循环;之后50 ℃加热到90 ℃(每6 s升高1 ℃)。建立熔解曲线,利用Bio-Rad CFX 2.1软件收集目的基因与内参基因的Ct值[13],通过qPCR package和Ct值计算每个技术重复的扩增效率,每个生物学样本包括3~5个技术重复,并参考Qiang等[14]的方法计算各个目的基因的相对表达量,具体方法如下:将Vg、Trf及β-actin扩增获得的Ct值绘出标准曲线,根据标准曲线所获得的线性计算公式,将样品的Ct值代入公式便得到目的基因的相对表达量。计算公式如下:
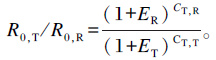
|
式中:ER为内参基因扩增效率;ET为目的基因扩增效率;CT, R为内参基因Ct值;CT, T为目的基因Ct值。
|
|
表 2 荧光定量PCR引物序列 Table 2 Primer sequences used in real time quantitative PCR |
试验数据用SPSS 17.0软件中的ANOVA程序进行差异显著性比较分析。
2 结果 2.1 饲粮粗蛋白质水平对培育蜂王初生重及胸宽的影响由表 3可知,蜂王的初生重随饲粮粗蛋白质水平的增加而升高,Ⅲ组和Ⅱ组的平均体重显著高于Ⅰ组(P<0.05) ,且Ⅲ组也显著高于Ⅱ组(P<0.05) 。蜂王的胸宽随饲粮粗蛋白质水平的增加而增加,但试验组之间差异不显著(P>0.05) 。
2.2 饲粮粗蛋白质水平对培育蜂王卵巢Vg、Trf相对表达量的影响由图 1可知,Ⅲ组和Ⅱ组培育蜂王的卵巢Vg相对表达量显著高于Ⅰ组(P<0.05) ,但Ⅲ组与Ⅱ组之间差异不显著(P>0.05) 。
|
|
表 3 饲粮粗蛋白质水平对培育蜂王初生重及胸宽的影响 Table 3 Effects of dietary crude protein level on birth weight and chest width of rearing queens |

|
数据柱标注相同字母表示差异不显著(P>0.05) ,不同字母表示差异显著(P<0.05) 。下图同。 Date columns with the same letters mean no significant difference (P>0.05) , while with different letters mean significant difference (P<0.05) . The same as below. 图 1 饲粮粗蛋白质水平对培育蜂王卵巢Vg基因表达量的影响 Figure 1 Effects of dietary protein level on the relative expression level of Vg in ovary of rearing queens |
由图 2可知,Ⅲ组和Ⅱ组培育蜂王的卵巢Trf相对表达量显著高于Ⅰ组(P<0.05) ,且Ⅲ组也显著高于Ⅱ组(P<0.05) 。

|
图 2 饲粮粗蛋白质水平对培育蜂王卵巢Trf相对表达量的影响 Figure 2 Effects of dietary protein level on the relative expression level of Trf in ovary of rearing queens |
营养是影响蜜蜂健康的关键因素,合理、均衡的营养物质供应以及适宜的环境是维持蜜蜂健康,保持高水平生产性能的重要条件。而蜂王是蜜蜂蜂群当中的主要繁殖个体,蜂王的质量决定蜂群群势的强弱。蜂王的初生重与蜂王的质量成正比关系,初生重大的蜂王,具有发育良好的卵巢管和数量较多的卵小管,这种蜂王具有较强的产卵力[15]。许少玉等[16]发现,中蜂蜂王初生重和卵小管数之间存在显著的线性关系,相关系数高达0.919。因此,蜂王初生重是衡量蜂王质量的一项重要指标。本研究发现:随着饲粮粗蛋白质水平的增加,中华蜜蜂蜂王初生重明显增加,Ⅲ组和Ⅱ组培育蜂王的初生重显著高于Ⅰ组,且Ⅲ组培育蜂王的初生重也显著高于Ⅱ组。其主要原因可能是因为:蛋白质是生命活动的物质基础,是生物体细胞的重要组成成分,为蜜蜂机体组织发育、更新、修补提供原料[17]。氨基酸是蛋白质的基本组成单位,其分解中间产物为乙酰辅酶A,而乙酰辅酶A是三大营养物质代谢的中间产物,它是与生命及与各种形式的生命活动紧密联系在一起的物质。在蜂王培育过程中饲喂高粗蛋白质水平的饲粮,使蜂王发育得到足够的蛋白质供应,提高营养物质之间相互调节、相互转换效率,使得机体能够充分发育,从而提高了蜂王的质量。Tarpy等[18]研究发现,胸宽与卵巢的发育存在一定的相关性,蜂王胸宽与卵巢管数存在正相关,胸宽的蜂王具有更多的卵巢管数,因此胸宽也是评估蜂王质量的重要指标,本研究发现:中华蜜蜂蜂王的胸宽随着饲粮蛋白质水平的增加而增加,但各试验组之间差异不显著,具体原因有待于进一步研究与分析。
Vg是一类大分子质量的糖脂复合蛋白质,广泛存在于卵生脊椎动物和非脊椎动物的血淋巴脂肪体和卵等器官中,是蜜蜂卵黄发生的关键物质,与蜂王的繁殖力、寿命和蜜蜂行为等生物学功能变化密切相关,卵黄蛋白原是卵黄蛋白的前体,卵黄蛋白为胚胎发育提供蛋白质、必需氨基酸等多种营养物质,卵巢发育好的蜂王其Vg具有较高的表达量[19-27],因此Vg的表达量也是衡量蜂王质量的重要指标。本研究发现:Ⅲ组和Ⅱ组卵巢Vg的相对表达量显著高于Ⅰ组,但Ⅲ组与Ⅱ组之间差异不显著。这也说明高蛋白质饲粮比低蛋白质饲粮培育的蜂王卵巢发育程度高。Human等[28]研究发现,高蛋白质饲粮会促进工蜂的卵巢发育。本试验结果与Human等[28]的试验结果很相似,这也说明饲粮蛋白质水平对蜂王质量存在影响,过低的蛋白质水平不利于蜂王卵巢发育。Ⅲ组与Ⅱ组卵巢Vg相对表达量差异不显著,可能是因为当饲粮中的蛋白质到达一定水平时已能满足卵巢发育所需的蛋白质的量。
Trf是蜜蜂体内铁的运输者,通过Trf受体介导的“结合铁的转铁蛋白-转铁蛋白受体”复合物的内吞,Trf将其结合的铁送入细胞内[29-30]。蜜蜂体内的Trf和铁结合蛋白能够调控游离Fe2+和Fe3+的含量,增强蜜蜂对感染寄生虫病的抵抗能力,降低因铁含量过高而引起的氧化反应对蜜蜂机体的危害[29, 31-33]。Koywiwattrakul等[34]研究发现,Trf在卵巢发育的工蜂上的表达量显著高于卵巢未发育的工蜂,Trf表达量的上调将通过一系列下游反应使卵巢活化,刺激工蜂的产卵行为。因此,Trf的表达量也可以为评价蜂王质量做参考。本研究发现:随着饲粮蛋白质水平的增加,Ⅲ组和Ⅱ组卵巢Trf相对表达量显著高于Ⅰ组,且Ⅲ组也显著高于Ⅱ组,说明给蜜蜂饲喂高蛋白质饲粮能够明显提升卵巢Trf含量,促进卵巢的发育,提高培育蜂王的抵抗力,促进卵巢的活化和产卵力。
4 结论① 饲粮粗蛋白质水平对中华蜜蜂育王质量存在影响,高蛋白质饲粮可以增加蜂王的初生重,促进蜂王卵巢Vg和Trf的表达。
② 育王时,中华蜜蜂饲粮粗蛋白质水平不应低于25%。
| [1] |
吴小波, 王子龙, 石元元, 等. 婚飞对中华蜜蜂性成熟处女蜂王sRNAs表达的影响[J].
吴小波, 王子龙, 石元元, 等. 婚飞对中华蜜蜂性成熟处女蜂王sRNAs表达的影响[J]. 中国农业科学, 2013 , 46 (17) :3721 –3728.
( 0) 0)
|
| [2] |
HAYDAK M H. Bee nutrition and pollen substitutes[J]. Apiacta, 1967 , 1 : 3 –8.
(0)
|
| [3] |
HAYDAK M H. Honey bee nutrition[J]. Annual Review of Entomology, 1970 , 15 : 143 –156.
DOI: 10.1146/annurev.en.15.010170.001043 (0)
|
| [4] |
KELLER I, FLURI P, IMDORF A. Pollen nutrition and colony development in honey bees:part Ⅰ.[J]. Bee World, 2005 , 86 (1) : 3 –10.
DOI: 10.1080/0005772X.2005.11099641 (0)
|
| [5] |
王改英, 吴在富, 杨维仁, 等. 饲粮蛋白质水平对意大利蜜蜂咽下腺发育及产浆量的影响[J].
王改英, 吴在富, 杨维仁, 等. 饲粮蛋白质水平对意大利蜜蜂咽下腺发育及产浆量的影响[J]. 动物营养学报, 2011 , 23 (7) :1147 –1152.
(0)
|
| [6] |
王改英, 李振, 杨维仁, 等. 日粮蛋白质水平对浙农大1号意大利蜜蜂产浆及咽下腺发育的影响[J].
王改英, 李振, 杨维仁, 等. 日粮蛋白质水平对浙农大1号意大利蜜蜂产浆及咽下腺发育的影响[J]. 江西农业大学学报, 2011 , 33 (6) :1176 –1180.
(0)
|
| [7] |
李成成, 杨维仁, 胥保华, 等. 意大利蜜蜂生长发育适宜蛋白供给水平及其对幼虫抗氧化活性的影响[J].
李成成, 杨维仁, 胥保华, 等. 意大利蜜蜂生长发育适宜蛋白供给水平及其对幼虫抗氧化活性的影响[J]. 中国农业科学, 2011 , 44 (22) :4714 –4720.
(0)
|
| [8] |
王改英, 杨维仁, 胥保华. 饲粮蛋白质水平对蜂群繁殖性能的影响[J].
王改英, 杨维仁, 胥保华. 饲粮蛋白质水平对蜂群繁殖性能的影响[J]. 应用昆虫学报, 2012 , 49 (2) :486 –489.
(0)
|
| [9] |
刘俊峰, 吴小波, 颜伟玉, 等. 饲粮蛋白水平对中华蜜蜂春繁性能及幼虫抗氧化性能的影响[J].
刘俊峰, 吴小波, 颜伟玉, 等. 饲粮蛋白水平对中华蜜蜂春繁性能及幼虫抗氧化性能的影响[J]. 江西农业大学学报, 2011 , 33 (5) :960 –964.
(0)
|
| [10] |
曾志将.
养蜂学[M]. 2版.北京: 中国农业出版社, 2009 : 101 -104.
(0)
|
| [11] |
秦秋红.东方蜜蜂与西方蜜蜂学习记忆比较及蜜蜂学习记忆相关分子机理分析[D].硕士学位论文.南昌:江西农业大学,2013.
(0)
|
| [12] |
WANG Z L, LIU T T, HUANG Z Y, et al. Transcriptome analysis of the Asian honey bee Apis cerana cerana[J]. PLoS One, 2012 , 7 (10) : e47954 .
DOI: 10.1371/journal.pone.0047954 (0)
|
| [13] |
WANG W X, TIAN L Q, HUANG Q, et al. Effects of 10-hydroxy-2-decenoic acid on the development of honey bee (Apis mellifera) larvae[J]. Journal of Apicultural Research, 2014 , 53 (1) : 171 –176.
DOI: 10.3896/IBRA.1.53.1.19 (0)
|
| [14] |
QIANG H, KRYGER P, CONTE Y L, et al. Survival and immune response of drones of a Nosemosis tolerant honey bee strain towards N. ceranae infections[J]. Journal of Invertebrate Pathology, 2012 , 109 (3) : 297 –302.
DOI: 10.1016/j.jip.2012.01.004 (0)
|
| [15] |
谢代癸. 对蜂王初生体重与质量的研究[J].
谢代癸. 对蜂王初生体重与质量的研究[J]. 中国蜂业, 1983 , 33 (2) :13 –14.
(0)
|
| [16] |
许少玉, 肖洪良, 李桂仙. 中蜂蜂王王台容积、初生重与卵小管数相关性的研究[J].
许少玉, 肖洪良, 李桂仙. 中蜂蜂王王台容积、初生重与卵小管数相关性的研究[J]. 中国蜂业, 1985 , 35 (1) :10 –11.
(0)
|
| [17] |
王颖, 马兰婷, 胥保华. 蜜蜂营养需要研究的必要性及策略[J].
王颖, 马兰婷, 胥保华. 蜜蜂营养需要研究的必要性及策略[J]. 动物营养学报, 2011 , 23 (8) :1269 –1272.
(0)
|
| [18] |
TARPY D R, KELLER J J, CAREN J R, et al. Assessing the mating ‘health’ of commercial honey bee queens.[J]. Journal of Economic Entomology, 2012 , 105 (1) : 20 –25.
DOI: 10.1603/EC11276 (0)
|
| [19] |
王秀秀, 杨明华, 李昌, 等. 东方蜜蜂卵黄原蛋白基因cDNA克隆及其基本生物信息学特征[J].
王秀秀, 杨明华, 李昌, 等. 东方蜜蜂卵黄原蛋白基因cDNA克隆及其基本生物信息学特征[J]. 中国蜂业, 2015 , 66 (3) :12 –15.
(0)
|
| [20] |
BLANK S, SEISMANN H, MCINTYRE M, et al. Vitellogenins are new high molecular weight components and allergens (Api m 12 and Ves v 6) of Apis mellifera and Vespula vulgaris venom[J]. PLoS One, 2013 , 8 (4) : e62009 .
DOI: 10.1371/journal.pone.0062009 (0)
|
| [21] |
WANG Y, BRENT C S, FENNERN E, et al. Gustatory perception and fat body energy metabolism are jointly affected by vitellogenin and juvenile hormone in honey bees[J]. PLoS Genetics, 2012 , 8 (6) : e1002779 .
DOI: 10.1371/journal.pgen.1002779 (0)
|
| [22] |
KAPHEIM K M, SMITH A R, IHLE K E, et al. Physiological variation as a mechanism for developmental caste-biasing in a facultatively eusocial sweat bee.[J]. Proceedings of the Royal Society B:Biological Sciences, 2012 , 279 (1732) : 1437 –1446.
DOI: 10.1098/rspb.2011.1652 (0)
|
| [23] |
CARDOEN R, ERNST U R, VAN VAERENBERGH M, et al. Differential proteomics in dequeened honeybee colonies reveals lower viral load in hemolymph of fertile worker bees[J]. PLoS One, 2011 , 6 (6) : e20043 .
DOI: 10.1371/journal.pone.0020043 (0)
|
| [24] |
GAUTHIER L, RAVALLEC M, TOURNAIRE M, et al. Viruses associated with ovarian degeneration in Apis mellifera L.Queens[J]. PLoS One, 2011 , 6 (1) : e16217 .
DOI: 10.1371/journal.pone.0016217 (0)
|
| [25] |
AMDAM G V, PAGE R E, J r. The developmental genetics and physiology of honeybee societies[J]. Animal Behaviour, 2010 , 79 (5) : 973 –980.
DOI: 10.1016/j.anbehav.2010.02.007 (0)
|
| [26] |
NUNES F M F, SIMÕES Z L P. A non-invasive method for silencing gene transcription in honeybees maintained under natural conditions[J]. Insect Biochemistry and Molecular Biology, 2009 , 39 (2) : 157 –160.
DOI: 10.1016/j.ibmb.2008.10.011 (0)
|
| [27] |
WHEELER D E, KAWOOYA J K. Purification and characterization of honey bee vitellogenin[J]. Archives of Insect Biochemistry and Physiology, 1990 , 14 (4) : 253 –267.
DOI: 10.1002/(ISSN)1520-6327 (0)
|
| [28] |
HUMAN H, NICOLSON S W, STRAUSS K, et al. Influence of pollen quality on ovarian development in honeybee workers (Apis mellifera scutellata)[J]. Journal of Insect Physiology, 2007 , 53 (7) : 649 –655.
DOI: 10.1016/j.jinsphys.2007.04.002 (0)
|
| [29] |
计成.
动物营养学[M]. 北京: 高等教育出版社, 2008 : 137 -142.
(0)
|
| [30] |
COWAN J A. Structural and catalytic chemistry of magnesium-dependent enzymes[J]. Biometals, 2002 , 15 (3) : 225 –235.
DOI: 10.1023/A:1016022730880 (0)
|
| [31] |
NICHOL H, LAW J H, WINZERLING J J. Iron metabolism in insects[J]. Annual Review of Entomology, 2002 , 47 (1) : 535 –559.
DOI: 10.1146/annurev.ento.47.091201.145237 (0)
|
| [32] |
DE SILVA D M, ASKWITH C C, KAPLAN J. Molecular mechanisms of iron uptake in eukaryotes[J]. Physiological Reviews, 1996 , 76 (1) : 31 –47.
(0)
|
| [33] |
KUCHARSKI R, MALESZKA R. Transcriptional profiling reveals multifunctional roles for transferrin in the honeybee,Apis mellifera[J]. Journal of Insect Science, 2003 , 3 (27) : 1 –8.
DOI: 10.1673/031.003.2701 (0)
|
| [34] |
KOYWIWATTRAKUL P, SITTIPRANEED P. Expression of vitellogenin and transferrin in activated ovaries of worker honey bees,Apis mellifera[J]. Biochemical Genetics, 2009 , 47 (1/2) : 19 –26.
(0)
|