




2. 华中农业大学动物科学技术学院、动物医学院, 武汉 430070
2. College of Animal Science and Animal Veterinary, Huazhong Agricultural University, Wuhan 430070, China
钠离子和氯离子是机体细胞外液的主要阳离子和阴离子,是维持细胞外液正常渗透压及容积的决定性因素,能调节体液的酸碱平衡,对营养物质的消化吸收具有重要作用。近年来,动物性原料在家禽配合饲粮中的含量不断下降,而钠、氯在植物性饲粮中含量极少,为了满足动物钠和氯的需要量,食盐常作为有效的矿物质补充料[1]。有研究表明,随着饲粮钠、氯水平的提高,肉鸡平均日采食量、饲料转化率、体增重显著升高,排泄物含水率也发生显著变化[2-5]。提高饲粮钠、氯水平显著影响血清阴、阳离子含量,使得血清渗透压显著升高[6],钠-葡萄糖共转运载体1(SGLT1) 表达也显著增加[7-8],而钠/氢交换载体 (NHE)2表达和钠-钾ATP (Na+-K+-ATP) 酶活性却显著降低[9-10],并且营养物质代谢水平也受到显著影响[6, 11]。目前,有关肉鸡钠、氯需要量的研究主要集中于生长性能和血清钠、钾、氯含量等指标方面,而关于其对鸡食欲调控、机体养分代谢及钠相关转运载体表达的影响尚缺乏研究。国内外尚无黄羽肉鸡钠、氯需要量的研究报道,实际生产上,黄羽肉鸡饲粮钠、氯添加水平多参考白羽肉鸡饲养标准 (NRC) 和生产经验。为此,为保障我国自主研发的黄羽肉鸡高效、标准化生产,迫切需要尽快确定黄羽肉鸡钠、氯需要量。本试验旨在研究饲粮钠、氯水平对黄羽肉鸡生长性能、排泄物含水率、血清生化指标和内分泌指标、肠道SGLT1和NHE2 mRNA表达量以及Na+-K+-ATP酶活性的影响,探讨1~21日龄黄羽肉鸡钠和氯的需要量及其作用机理,为黄羽肉鸡饲粮配制提供科学依据。
1 材料与方法 1.1 试验动物与分组选用1日龄健康、发育良好的快大型岭南黄羽肉公鸡共1 200只,根据体重随机分成5组,每组6个重复,每个重复40只鸡。试验鸡分组遵循各组间和重复间体重无显著差异 (P>0.05) 的原则。
1.2 试验设计与饲粮处理试验采用单因子随机分组设计,对照组 (1组) 采用玉米-豆粕型基础饲粮;其余各组参考《鸡饲养标准》(NY/T 33-2004) 和NRC (1994) 设计饲粮钠、氯含量比为1 : 1,并通过食盐与碳酸氢钠等量取代部分沸石粉使饲粮钠和氯含量均分别为0.1%、0.2%、0.3%和0.4%。除钠、氯水平外,各组饲粮营养水平均参考《鸡饲养标准》(NY/T 33-2004)。各组饲粮组成及营养成分见表 1。
|
|
表 1 饲粮组成及营养水平 (风干基础) Table 1 Composition and nutrient levels of diets (air-dry basis) |
饲养试验于2015年7月在广东省农业科学院动物科学研究所动物营养研究室试验场进行,采用地面平养,地面铺放稻壳,各组饲养环境条件一致。试鸡自由采食、饮水,按照常规饲养操作规程和免疫程序进行饲养和免疫。每天08:00、14:30和20:00测定鸡舍的温度和相对湿度,3次测定的平均值作为当天鸡舍的平均温度和相对湿度[试验期间鸡舍温度和相对湿度分别为 (31.04±1.19)℃、(81.92±9.93)%]。
1.4 样品的采集与处理试鸡21日龄时,每重复挑选2只健康、接近平均体重的试鸡,称重并记录后,翅静脉采血于非抗凝管中,静置30 min后,3 500 r/min离心10 min,制备血清,分装至500 μL的EP管中,-80 ℃冻存,待测血清生化指标和内分泌指标;试鸡放血屠宰后剖摘腺胃和肌胃,测定腺胃内pH;试验1、3和5组完整分离回肠和十二指肠,沿纵向剖开,用冷却的磷酸缓冲盐水冲洗,吸水纸吸干,取肠段约1 cm于离心管中,立即放入液氮中,然后转至-80 ℃低温冰箱中保存,用于测定SGLT1和NHE2的mRNA表达量和Na+-K+-ATP酶活性。
1.5 测定指标与方法 1.5.1 生长性能试验过程中,一旦出现死鸡,立即称死鸡重和剩料量,以消除死鸡对最后试验结果的影响,记录死鸡数量,同时应立即查明原因,采取适当措施。各阶段饲养试验结束前1天20:00断料供水,次日08:00以重复为单位称重,并统计耗料量,计算各生长阶的平均日采食量、平均日增重和料重比,同时还计算死亡率。
1.5.2 饮水量每天08:30记录前1 d的剩水重量和当天的加水重量,并用蒸发量对其进行校正 (2个装有已知体积水的水槽,其减少的重量即蒸发量),加水重量减去剩水重量就是1 d的饮水量。
1.5.3 排泄物含水率试验结束前5天,每重复选接近平均体重试鸡2只置于笼中进行排泄物收集。收集的排泄物先称重,然后于烘箱中 (60 ℃) 烘至恒重。
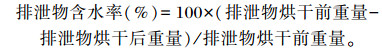
|
将pH计 (DELTA320,梅特勒-托利多) 探针插入待测腺胃,测定腺胃上部、中部和下部pH,取其平均值。pH计使用前需要用pH 4.01、6.86和9.18的标准缓冲液进行校正。
1.5.5 血清生化、内分泌指标血清钠、钾、氯、尿酸、葡萄糖、总胆固醇及甘油三酯含量测定使用南京建成生物工程研究所所产试剂盒,所用仪器为全波长酶标仪 (Bio-Rad 550,美国)。血清胆囊收缩素、胃饥饿素和生长激素含量采用酶联免疫分析法 (ELISA) 测定,试剂盒购于北京方程生物科技有限公司,所用仪器为全波长酶标仪。
血清的渗透压=2×钠+2×钾+葡萄糖+尿酸[12]。
1.5.6 十二指肠Na+-K+-ATP酶活性准确称取十二指肠约0.1 g,按重量 (g):体积 (mL)=1 : 9的比例,加入9倍体积的生理盐水,冰水浴条件下机械匀浆,2 500 r/min离心10 min,取上清液,再用生理盐水10倍稀释成1%,用二甲酸喹啉 (BCA) 蛋白质定量分析试剂盒 (购于美国Thermo Fisher Scientific公司) 测定总蛋白含量,用超微量Na+-K+-ATP酶测试盒 (购于南京建成生物工程研究所) 检测十二指肠Na+-K+-ATP酶活性。Na+-K+-ATP酶活性定义:规定每小时每毫克组织蛋白质的组织中ATP酶分解ATP产生1 μmol无机磷的量为1个ATP酶活性单位。
1.5.7 回肠SGLT1和十二指肠NHE2的mRNA表达量测定采用Trizol一步法提取组织样的总RNA,按照TaKaRa公司的反转录试剂盒PrimeScriptTM RT reagent Kit (RR047A) 进行反转录,根据NCBI中GenBank提供的基因序列,使用Prime Premier 5.0软件对SGLT1、NHE2 2个基因和1个内参基因β-肌动蛋白 (β-actin) 进行引物序列设计,引物由上海生工生物工程有限公司广州分公司合成。设计的引物序列如表 2所示。以鸡回肠和十二指肠cDNA为模板,依照表 2中相应的引物,将各成分 (反应体系为2 μL cDNA、1 μL Forward primer、1 μL Reverse primer、10 μL SYBR Green PCR Master Mix和6 μL ddH2O) 加到实时荧光定量PCR专用96孔板中。反应程序为:95 ℃预变性1 min;95 ℃变性15 s,在基因特异性退火温度条件下退火15 s,72 ℃延伸40 s,40个循环。以β-actin作为内参,对获得的信号、数据进行处理,在确定各目的基因与内参基因扩增效率接近的基础上,采用△△Ct法来度量各基因的mRNA表达丰度,每个样品的平均Ct值减去对应样品的β-actin的平均Ct值,得到△Ct值,然后再以其中1个样本为参照,将每个△Ct值减去该样品的△Ct值即得到△△Ct值,最后计算平均相对表达量为2-△△Ct。
|
|
表 2 实时荧光定量PCR引物序列及参数 Table 2 Sequences and parameters of primers for the real-time PCR |
试验数据采用SAS V8.0软件中的GLM程序进行方差分析,并进行Duncan氏多重比较,统计显著水平为P < 0.05。当数据方差分析和Duncan氏多重比较达到显著水平时,再根据Robbins等[13]的研究进行回归分析,二次曲线的最高点对应的横坐标值的95%即为黄羽肉鸡钠、氯营养需要量。
2 结果与分析 2.1 饲粮钠、氯水平对黄羽肉鸡生长性能、饮水量与排泄物含水率的影响 2.1.1 饲粮钠、氯水平对黄羽肉鸡生长性能的影响由表 3可以看出,饲粮钠、氯水平显著影响试鸡平均日采食量、平均日增重、料重比和死亡率 (P < 0.05)。对照组平均日采食量和平均日增重显著低于其他各组 (P < 0.05),料重比显著高于其他各组 (P < 0.05)。饲粮钠、氯水平均为0.2%时,平均日采食量和平均日增重达到最大值,显著高于对照组和0.1%钠、氯水平组 (P < 0.05),与0.4%钠、氯水平组无显著差异 (P>0.05)。钠、氯水平均高于0.1%的各组间料重比无显著差异 (P>0.05)。对照组死亡率显著高于除0.4%钠、氯水平组的其他3组 (P < 0.05)。综合上述结果,在本试验条件下,1~21日龄黄羽肉鸡饲粮钠、氯水平均为0.2%便可获得最佳生长性能,因此饲粮钠、氯适宜水平均为0.2%。
|
|
表 3 饲粮钠、氯水平对1~21日龄黄羽肉鸡生长性能、饮水量、排泄物含水率、死亡率及腺胃pH的影响 Table 3 Effects of dietary sodium and chlorine levels on growth performance, water consumption, excreta moisture rate, mortality and proventriculus pH in yellow-feathered broilers of 1 to 21 days of age |
参考Robbins等[13]的研究,通过非线性回归模型建立饲粮钠、氯水平与平均日增重间的二次折线回归方程,其中x表示饲粮钠、氯水平,y表示平均日增重 (图 1),得出:
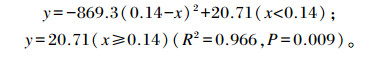
|

|
图 1 饲粮钠、氯水平对1~21日龄黄羽肉鸡平均日增重的影响 Figure 1 Effects of dietary sodium and chlorine levels on average daily gain of yellow-feathered broilers of 1 to 21 days of age |
该方程表明饲粮钠、氯水平均为0.14%时试鸡体增重达到平台期,即该模型估测得到的饲粮钠、氯需要量为0.14%。可见,由非线性回归模型估测得到的需要量低于试验直观评定的结果 (0.2%)。
2.1.2 饲粮钠、氯水平对黄羽肉鸡饮水量与排泄物含水率的影响由表 3可以看出,随着饲粮钠、氯水平的提高,试鸡平均日饮水量显著提高 (P < 0.05),当钠、氯水平为0.2%~0.3%时,平均日饮水量趋于平缓。从数值上可以看出,排泄物含水率与饲粮钠、氯水平呈二次曲线变化 (P=0.046),钠、氯水平均为0.3%时达到最高点,并且各试验组都显著高于对照组 (P < 0.05)。
参考Robbins等[13]的研究,通过非线性回归模型建立饲粮钠、氯水平与排泄物含水率间的二次曲线回归方程,其中x表示饲粮钠、氯水平,y表示排泄物含水率 (图 2),得出:
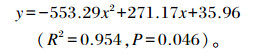
|

|
图 2 饲粮钠、氯水平对黄羽肉鸡排泄物含水率的影响 Figure 2 Effects of dietary sodium and chlorine levels on excreta moisture rate of yellow-feathered broilers |
该二次曲线的最高点对应的横坐标值的95%为0.23%,即该模型估测饲粮钠、氯含量均为0.23%时排泄物含水率最大,略低于试验直观评定的结果 (0.3%)。
2.2 饲粮钠、氯水平对黄羽肉鸡腺胃pH的影响由表 3可以看出,随着饲粮钠、氯水平的提高,试鸡腺胃pH并未发生显著变化 (P>0.05)。
2.3 饲粮钠、氯水平对黄羽肉鸡血清指标的影响 2.3.1 饲粮钠、氯水平对黄羽肉鸡血清钠、钾、氯含量和渗透压的影响由表 4可以看出,血清钠、钾含量和渗透压分别随饲粮钠、氯水平的提高呈二次折线变化。对照组血清钠含量和渗透压均显著低于其他组 (P < 0.05),而血清钾含量则显著高于其他组 (P < 0.05)。饲粮钠、氯水平在0.1%~0.4%内,随着饲粮钠、氯水平的提高,血清钠、钾含量及渗透压均无显著变化 (P>0.05)。从数值上看,饲粮钠、氯水平均为0.2%时,血清氯含量达到最高值,显著高于对照组 (P < 0.05),而与其他组间无显著差异 (P>0.05)。综合上述结果,在本试验条件下,根据血清钠、钾、氯含量和渗透压值直观判定黄羽肉鸡饲粮钠、氯适宜水平均为0.1%,但通过对血清钠、钾和渗透压进行非线性回归分析估测得到对应的钠、氯需要量均为0.13%、0.18%和0.13%。
|
|
表 4 饲粮钠、氯水平对21日龄黄羽肉鸡血清生化指标和内分泌指标的影响 Table 4 Effects of dietary sodium and chlorine levels on serum biochemical and endocrine parameters of yellow-feathered broilers at 21 days of age |
由表 4可以看出,血清尿酸、葡萄糖和甘油三酯含量分别随饲粮钠、氯水平的提高呈二次折线降低。当饲粮钠、氯水平均为0.1%时,试鸡血清尿酸、葡萄糖和甘油三酯含量都显著低于对照组 (P < 0.05),而与其他组并没有显著差异 (P>0.05)。饲粮钠、氯水平均为0.1%时,血清总胆固醇含量最低,显著低于对照组以及0.3%和0.4%钠、氯水平组 (P < 0.05),与0.2%钠、氯水平组没有显著差异 (P>0.05)。综合上述结果,在本试验条件下,根据血清尿酸、葡萄糖和甘油三酯含量直观判定黄羽肉鸡饲粮钠、氯适宜水平均为0.1%,但通过对血清尿酸、葡萄糖和甘油三酯含量进行非线性回归分析估测得到钠、氯需要量均分别为0.13%、0.21%和0.10%。
2.3.3 饲粮钠、氯水平对黄羽肉鸡血清胆囊收缩素、胃饥饿素和生长激素含量的影响由表 4可以看出,饲粮钠、氯水平对试鸡血清胆囊收缩素、胃饥饿素和生长激素含量无显著影响 (P>0.05),但随着饲粮钠、氯水平的提高,血清胆囊收缩素含量呈缓慢下降的趋势,而血清胃饥饿素含量呈缓慢升高的趋势。
2.4 饲粮钠、氯水平对黄羽肉鸡小肠SGLT1、NHE2 mRNA表达量和Na+-K+-ATP酶活性的影响由表 5可以看出,随着饲粮钠、氯水平的提高,试鸡回肠SGLT1 mRNA表达量先升高后下降,饲粮钠、氯水平均为0.2%时达到最大值,且显著高于其他2组 (P < 0.05)。十二指肠NHE2 mRNA表达量和Na+-K+-ATP酶活性呈下降趋势,且对照组显著高于其他2组 (P < 0.05)。
|
|
表 5 饲粮钠、氯水平对21日龄黄羽肉鸡小肠SGLT1、NHE2 mRNA表达量和Na+-K+-ATP酶活性的影响 Table 5 Effects of dietary sodium and chlorine levels on the expression of SGLT1 and NHE2 mRNA and the activity of Na+-K+-ATPase in small intestine of yellow-feathered broilers at 21 days of age |
本试验结果表明,随着饲粮钠、氯水平的提高,1~21日龄试鸡平均日采食量与平均日增重显著提高,在饲粮钠、氯水平均为0.2%时达到平台期,料重比则随着饲粮钠、氯水平的提高而显著降低,在饲粮钠、氯水平均为0.1%时达到平台期。这与Murakami等[2]的结论相似,平均日增重分别随着饲粮钠和氯水平的提高而提高,在饲粮钠、氯分别提高到0.25%和0.20时达到平台期。Oviedo-Rondón等[14]也得出类似结论,1~21日龄肉鸡生长性能均随饲粮钠和氯水平呈二次曲线变化,最佳饲粮钠、氯水平分别是0.28%和0.25%。Vieira等[15]研究表明,饲粮钠水平为0.48%时肉鸡死亡率显著高于其他处理。本试验条件下,饲粮添加0.1%、0.2%和0.3%钠和氯,试鸡死淘率降低了4个百分点以上。可见,饲粮适宜钠、氯水平能够显著改善肉鸡的生长性能。
钠离子和氯离子是机体细胞外液的主要阳离子和阴离子,是维持细胞外液正常渗透压及容积的决定性因素[16],钠、氯摄入后在体内转化成酸和碱,在过酸或过碱的环境下,动物机体的新陈代谢会更倾向于调节过量的酸或碱,从而会使生长发育受到影响[17]。随着钠、氯水平的提高,血清钠、氯含量及渗透压显著升高,而血液pH并没有发生显著变化,但通过血液碳酸氢根、总二氧化碳和碱剩含量的变化可以看出,血液的缓冲能力逐渐提高[11, 18]。由此提示,饲粮钠、氯水平可以通过调整血液渗透压和血液缓冲能力影响生长性能。
3.1.2 饲粮钠、氯水平对黄羽肉鸡饮水量的影响本试验结果表明,随着饲粮钠、氯水平的提高,1~21日龄试鸡平均日饮水量显著提高,这与Vieira等[15]、Maiorka等[19]和Mushtaq等[4]的研究结果一致。随着钠、氯水平的提高,血清钠、氯含量及渗透压显著升高[11, 18],为维持正常的血清钠、氯及渗透压水平,饮水量便会提高,大量的饮水会稀释血清钠、氯含量及降低渗透压。
3.1.3 饲粮钠、氯水平对黄羽肉鸡排泄物含水率的影响关于饲粮钠、氯水平对排泄物含水率的影响争议较大。Murakami等[3]研究表明排泄物含水率随钠水平升高呈正线性关系,而与氯水平无关。Mushtaq等[4]则认为,排泄物含水率是由饲粮中的钠、氯共同影响的。Koreleski等[5]研究表明排泄物含水率与饲粮钾呈正相关关系,而与饲粮钠无显著关系。本试验结果表明1~21日龄试鸡排泄物含水率随饲粮钠、氯水平呈二次曲线变化,饲粮钠、氯水平为0.23%时达到最大值,这与本试验血清氯的变化趋势相似,但血清氯与水盐代谢间的确切关系尚需要进一步研究。
3.2 饲粮钠、氯水平对黄羽肉鸡腺胃pH的影响腺胃pH主要是由胃液中胃酸决定的,胃酸是由通过胃质子泵进入胃腔的氢离子与顶膜转运至胃腔的氯离子形成的,腺胃pH对胃内酶活性有重要作用[20]。关于饲粮钠、氯水平对腺胃pH影响并没有相关报道,本试验结果表明,饲粮钠、氯水平对腺胃pH无有显著影响。
3.3 饲粮钠、氯水平对黄羽肉鸡血清指标的影响 3.3.1 饲粮钠、氯水平对黄羽肉鸡血清生化指标的影响钠、钾、氯是维持血液渗透压的主要离子,在维持强离子差方面也有密切关系[20]。本试验结果表明,随着饲粮钠、氯水平的提高,试鸡血清钠、氯含量和渗透压都显著升高,血清钾含量显著降低,当饲粮钠、氯水达到0.1%时,试鸡血清钠、钾含量和渗透压便到达平台期。Mushtaq等[4]报道,肉鸡血清中钠含量随饲粮中氯添加量提高呈线性升高。鲍庆晗等[21]也表明,饲粮氯水平提高会引起蛋鸭血清钠含量的升高。以上试验证实,正常情况下细胞外液中钠离子、碳酸氢根离子、氯离子之间有一个较恒定的常数,即钠离子=碳酸氢根离子+氯离子+阴离子间隙,当血清氯含量升高时,血清钠含量相应增加,以保证阴阳离子总和相等[22]。本试验中饲粮钠水平是由食盐与碳酸氢钠提供,随着饲粮钠、氯水平的提高,碳酸氢根含量也由0提高到0.42%,且饲粮中钠、氯离子相等,因为电中性原理,当碳酸氢根高到一定程度必然会取代部分氯,数据表明在饲粮碳酸氢根为0.22%时开始起作用,即随着饲粮钠、氯水平的提高,血清氯先升高后下降。对于血清钾含量,本试验饲粮钾水平是固定的,当饲粮钠、氯添加水平为0时,饲粮中主要阳离子便是钾离子,随着饲粮钠、氯水平的提高,钠离子便成了主要的阳离子,根据电中性原理,血清钾含量必然会下降。
鲍庆晗[18]和王安等[11]研究表明,在低氯和高氯的情况下蛋鸭血清尿酸含量较高。本试验结果也发现,随着饲粮钠、氯水平的提高,试鸡血清尿酸含量降低,在饲粮钠、氯水平为0.1%时达到平台期。饲粮钠、氯水平较低时,试鸡血清尿酸含量很高,说明内源氮排出量增加,沉积受到影响,从而影响蛋白质代谢,破坏了氨基酸的平衡。徐奇友等[23]报道,血糖丰富时,胰岛素通过影响细胞膜的转运系统促进细胞对氨基酸和葡萄糖的摄取,增加蛋白质和糖原的合成。可见葡萄糖还是蛋白质合成的重要底物,血糖含量可以作为判断糖类代谢状况的指标,血糖含量高时表明糖类代谢紊乱,反之则表明血糖平衡。本试验结果表明,随着饲粮钠、氯水平的提高,试鸡血清葡萄糖和甘油三酯含量显著降低,总胆固醇含量呈先降低后升高的趋势,且当饲粮钠、氯水平均为0.1%时,试鸡血清葡萄糖、甘油三酯和总胆固醇含量均达到平台期。由此提示,在本试验条件下,饲粮低钠、氯水平可能通过调节血清尿酸、葡萄糖、甘油三酯和总胆固醇含量影响机体蛋白质、糖和脂类代谢,但饲粮钠、氯达到一定水平时,这一调控作用下降,也说明机体具有维持血清尿酸、葡萄糖、甘油三酯和总胆固醇含量稳定的能力。
综上所述,随着饲粮钠、氯水平的提高,血清钠、钾、氯、碳酸氢根相互调节,为了遵循阴阳离子平衡,同时维持渗透压稳定,当饲粮钠、氯水平升高到0.1%以后,血清渗透压趋于稳定。营养物质代谢水平也受饲粮钠、氯水平影响,其中葡萄糖代谢最为敏感。血清渗透压与生长性能及以葡萄糖为主的营养物质代谢水平的变化趋势相似,提示饲粮钠、氯水平可能通过调节血清渗透压来影响营养物质代谢水平,从而影响试鸡生长性能。
3.3.2 饲粮钠、氯水平对黄羽肉鸡血清胆囊收缩素、胃饥饿素和生长激素含量的影响食盐作为调味剂具有一定的咸味,在具有咸味的盐类物质中,其咸度也是很高的,其咸味也随着浓度的提高而提高,其在食欲调控中有很大的作用[24]。胆囊收缩素是一种由胃肠道黏膜Ⅰ细胞分泌的多肽激素,具有抑制摄食行为、影响激素分泌等效应[25]。胆囊收缩素主动免疫和被动免疫都可以提高动物的采食量和增重[26-27]。胃饥饿素是胃内产生的一种肽,是生长激素促分泌激素受体1A的一种内源性配体,它能够调节食欲、进食和身体构成[28]。Gnanapavan等[29]报道胃饥饿素还可以增强胃动素和胃泌素的功能,通过调节胃酸及消化酶的分泌增强胃肠蠕动等,来增加胃排空的速率,增加体重。本试验显示,随着饲粮钠、氯水平的提高,试鸡平均日采食量显著提高,但血清胆囊收缩素和胃饥饿素含量均未受显著影响。可见,在本试验条件下,饲粮钠、氯水平提高未通过胆囊收缩素和胃饥饿素调控试验鸡采食量,可能通过其他食欲调控因子引起试验鸡食欲增强,但其确切机制还需要进一步研究。本试验条件下,饲粮钠、氯水平对试鸡血清生长激素含量并没有显著影响。因此,饲粮钠、氯并没有通过调节生长激素的分泌来影响生长性能。
3.4 饲粮钠、氯水平对黄羽肉鸡小肠SGLT1、NHE2 mRNA表达量和Na+-K+-ATP酶活性的影响SGLT1是动物小肠吸收葡萄糖的主要决定因素。饲粮钠水平低并没有引起SGLT1 mRNA的变化[30]。Garriga等[31]利用蛋白质印迹法 (Western Blot) 检测发现,饲粮钠水平低会导致回肠和直肠SGLT1表达显著降低,而空肠中没有变化,而当饲粮钠水平提高时,SGLT1表达便会恢复[10]。可见,饲粮钠、氯水平对回肠和空肠SGLT1的表达是有影响的,这种影响被称为转录后效应[7-8]。低钠会引起继发性醛固酮症,并且所有观察到的影响被认为是有醛固酮引起的[32-33],所以醛固酮可能是饲粮钠水平影响SGLT1转录后效应的关键。与上述结果不同,本试验结果表明,随着饲粮钠、氯水平的提高,试鸡回肠SGLT1 mRNA表达量先升高后降低,饲粮钠、氯水平均为0.2%时达到最大值。这也可能与试验鸡日龄有关,上述各研究报道均以中、大鸡为试验对象,而本试验结果为21日龄鸡的,结果提示随着日龄的增长,机体可能对于基因转录表达调控能力增强,而对于转录后效应的调控能力并未增强。
NHE是一类重要的膜蛋白,其生理作用是进行细胞外钠和细胞内氢的电中性交换,在动物体细胞已发现的NHE有9种亚型。王修启等[34]研究表明,肉鸡NHE2 mRNA表达量在十二指肠最高。Donowitz等[9]研究表明,在低钠水平下,结肠NHE2 mRNA表达量显著高于高钠水平的。与上述结果相似,本试验条件下,随着饲粮钠、氯水平的提高,试鸡十二指肠NHE2 mRNA表达量显著下降。可见,饲粮钠、氯水平对NHE2 mRNA的表达有显著影响。
Na+-K+-ATP酶,又称为三磷酸腺苷酶,是一类能将ATP催化水解为ADP和磷酸根离子的酶,这是一个释放能量的反应,会为主动转运的物质 (钠离子、钙离子、氢离子、氯离子、葡萄糖等) 提供能量[35]。随着饲粮钠水平的提高,Na+-K+-ATP酶活性下降[7-10]。与上述结果相似,本试验结果表明,随着饲粮钠、氯水平的提高,试鸡十二指肠Na+-K+-ATP酶活性下降,可能会抑制肠道对钠离子、氯离子和氢离子的转运。
饲粮钠、氯水平对试鸡小肠SGLT1、NHE2 mRNA表达量和Na+-K+-ATP酶活性的影响是试鸡对饲粮钠、氯水平进行自主适应的一种生理上的表现,其影响的原理与内分泌系统有着重要的联系,可能是受到某种或某些激素 (醛固酮、生长激素、皮质醇、胰岛素样生长因子) 的控制[36]。随着饲粮钠、氯水平提高,SGLT1 mRNA与生长性能的变化趋势不一致,可见,随着饲粮钠、氯水平的变化,SGLT1参与了对生长性能的调节,但不是主要机制。随着饲粮钠、氯水平的提高,NHE2 mRNA表达量与Na+-K+-ATP酶活性显著降低,可能抑制钠离子的吸收与氢离子的分泌,由此提示,NHE2和Na+-K+-ATP酶可能是参与维持渗透压稳定与酸碱平衡的重要机制。
3.5 肉鸡钠、氯需要量的探讨畜禽养分需要量常采用计量反应法进行评定,即通过饲粮中添加不同梯度水平的养分,然后测定反映动物各种特性的敏感指标状况,如生长性能、血液指标及基因表达等,最后经综合判断来确定某一养分的营养需要量。但是,以不同的指标为判断标准得出的需要量并不一致。Oviedo-Rondón等[14]根据生长性能给出1~21日龄肉鸡最佳饲粮钠、氯水平分别为0.28%和0.23%,以血气分析为评定指标,当饲粮氯水平为0.19%时,血气分析指标最佳。本试验通过国内传统的营养需要量研究方法结合非线性回归模型估测得到饲粮钠、氯的需要量,以生长性能、血清渗透压和葡萄糖含量为标准得到的适宜饲粮钠、氯水平也存在差异。综合上述讨论,饲粮钠、氯水平通过调整血液渗透压与酸碱平衡来影响营养物质代谢 (血清葡萄糖最敏感),进而影响生长性能,因此以生长性能、血清渗透压和葡萄糖含量为标准来判断黄羽肉鸡钠、氯营养需要量 (表 6)。根据平均日增重主观判定和血清葡萄糖含量的回归估测得到的钠、氯需要量与NRC (1994) 肉用仔鸡的推荐量基本一致,但明显高于我国黄羽肉鸡饲养标准 (2004)。经平均日增重和血清渗透压回归估测得到的钠、氯需要量则明显低于NRC (1994)、Murakami等[2]和Oviedo-Rondón等[14]的推荐值。由此提示,肉鸡可能因品种、饲养方式和评判指标等不同使其钠、氯需要量推荐值存在差异,因此,在实际生产中,应根据实际情况选用恰当的标准进行科学饲粮配制。
|
|
表 6 不同评价指标得出的1~21日龄黄羽肉鸡钠、氯需要量 Table 6 Requirements of sodium and chlorine for yellow-feathered broilers from 1 to 21 days of age by different evaluation indices |
本试验条件下:
①饲粮补充钠、氯显著提高了试鸡平均日增重、平均日采食量、平均日饮水量和排泄物含水率,降低了死亡率,平均日增重随饲粮钠、氯水平的提高呈二次折线升高。
②饲粮补充钠、氯显著提高了血清渗透压,降低了血清尿酸、葡萄糖、总胆固醇、甘油三酯含量和回肠SGLT1 mRNA表达量,降低了十二指肠NHE2 mRNA表达量和Na+-K+-ATP酶活性。
③以生长性能为评定指标,通过非线性回归模型估测得到1~21日龄阶段快大型黄羽肉鸡饲粮适宜钠、氯水平均为0.14%。
| [1] | 于炎湖. 畜禽饲粮中应用食盐的技术问题[J]. 中国饲料, 2004(16) :3–5, 7. |
| [2] | MURAKAMI A E, SALEH E A, ENGLAND J A, et al. Effect of level and source of sodium on performance of male broilers to 56 days[J]. Journal of Applied Poultry Research, 1997, 6(2): 128–136. DOI: 10.1093/japr/6.2.128 |
| [3] | MURAKAMI A E, WATKINS S E, SALEH E A, et al. Estimation of the sodium and chloride requirements for the young broiler chick[J]. Journal of Applied Poultry Research, 1997, 6(2): 155–162. DOI: 10.1093/japr/6.2.155 |
| [4] | MUSHTAQ T, SARWAR M, NAWAZ H, et al. Effect and interactions of dietary sodium and chloride on broiler starter performance (hatching to twenty-eight days of age) under subtropical summer conditions[J]. Poultry Science, 2005, 84(11): 1716–1722. DOI: 10.1093/ps/84.11.1716 |
| [5] | KORELESKI J, ŚWIATKIEWICZ S, ARCZEWSKA A. The effect of dietary potassium and sodium on performance, carcass traits, and nitrogen balance and excreta moisture in broiler chicken[J]. Journal of Animal and Feed Sciences, 2010, 19(2): 244–256. DOI: 10.22358/jafs/66285/2010 |
| [6] | 赵琬婧, 王安. 钠和氯对生长蛋鸭生产性能和血液生化指标的影响[J]. 东北农业大学学报, 2008, 39(7) :66–71. |
| [7] | BARFULL A, GARRIGA C, MITJANS M, et al. Ontogenetic expression and regulation of Na+-D-glucose cotransporter in jejunum of domestic chicken[J]. American Journal of Physiology-Gastrointestinal and Liver Physiology, 2002, 282(3): G559–G564. DOI: 10.1152/ajpgi.00262.2001 |
| [8] | BARFULL A, GARRIGA C, TAULER A, et al. Regulation of SGLT1 expression in response to Na+ intake[J]. American Journal of Physiology:Regulatory, Integrative and Comparative Physiology, 2002b, 282(3): R738–R743. DOI: 10.1152/ajpregu.00263.2001 |
| [9] | DONOWITZ M, DE LA HORRA C, CALONGE M L, et al. In birds, NHE2 is major brush-border Na+/H+ exchanger in colon and is increased by a low-NaCl diet[J]. American Journal of Physiology:Regulatory, Integrative and Comparative Physiology, 1998, 274(6): R1659–R1669. |
| [10] | GARRIGA C, MORETÓ M, PLANAS J M. Effects of resalination on intestinal glucose transport in chickens adapted to low Na+ intakes[J]. Experimental Physiology, 2000, 85(4): 371–378. DOI: 10.1111/eph.2000.85.issue-4 |
| [11] | 王安, 李士平, 任延铭. 5~11周龄金定蛋鸭实用日粮中氯化钠添加量的研究[J]. 动物营养学报, 2010, 22(2) :460–465. |
| [12] | 张志斌. 物理化学教学与兴趣教育[J]. 大学化学, 2006, 21(5) :13–16. |
| [13] | ROBBINS K R, SAXTON A M, SOUTHERN L L. Estimation of nutrient requirements using broken-line regression analysis[J]. Journal of Animal Science, 2006, 84(13): E155–E165. |
| [14] | OVIEDO-RONDÓN E O, MURAKAMI A E, FURLAN A C, et al. Sodium and chloride requirements of young broiler chickens fed corn-soybean diets (one to twenty-one days of age)[J]. Poultry Science, 2001, 80(5): 592–598. DOI: 10.1093/ps/80.5.592 |
| [15] | VIEIRA S L, PENZ A M, POPHAL S, et al. Sodium requirements for the first seven days in broiler chicks[J]. The Journal of Applied Poultry Research, 2003, 12(3): 362–370. DOI: 10.1093/japr/12.3.362 |
| [16] | 冷向军, 王康宁, 杨凤, 等. 酸化剂对仔猪生长和体内酸碱平衡的影响[J]. 动物营养学报, 2003, 15(2) :49–53, 64. |
| [17] | MONGIN P. Recent advances in dietary anion-cation balance:applications in poultry[J]. Proceedings of the Nutrition Society, 1981, 40(3): 285–294. DOI: 10.1079/PNS19810045 |
| [18] | 鲍庆晗.氯对生长蛋鸭生产性能及血液生化指标的影响[D].硕士学位论文.哈尔滨:东北农业大学, 2008. |
| [19] | MAIORKA A, MAGRO N, BARTELS H A S, et al. Different sodium levels and electrolyte balances in pre-starter diets for broilers[J]. Revista Brasileira de Ciência Avícola, 2004, 6(3): 143–146. |
| [20] | 李鉴轩. 家畜生理学[M]. 西宁: 青海人民出版社, 1992: 142. |
| [21] | 鲍庆晗, 王安, 王洋. 氯对生长蛋鸭生产性能和体内酸碱平衡的影响[J]. 动物营养学报, 2007, 19(4) :321–326. |
| [22] | 韩志钧. 血气酸碱分析[M]. 2版. 沈阳: 辽宁科学技术出版社, 2004: 58-64. |
| [23] | 徐奇友, 单安山, 李建平. 乳铁蛋白对早期断奶仔猪血清生化指标影响的研究[J]. 中国畜牧杂志, 2006, 42(3) :34–36. |
| [24] | 吴海舟, 张迎阳, 唐静, 等. 降低肉制品中氯化钠含量研究进展[J]. 肉类研究, 2014, 28(6) :22–26. |
| [25] | 丁雪梅, 张克英, 陈代文. 胆囊收缩素主动免疫产蛋鸡的营养生理效应研究[J]. 畜牧兽医学报, 2005, 36(6) :564–568. |
| [26] | PEKAS J C. Effect of cholecystokinin immunization, enhanced food intake and growth of swine on lean yield and carcass composition[J]. The Journal of Nutrition, 1991, 121(4): 563–567. |
| [27] | REIDELBERGER R D, VARGA G, LIEHR R M, et al. Cholecystokinin suppresses food intake by a nonendocrine mechanism in rats[J]. American Journal of Physiology:Regulatory, Integrative and Comparative Physiology, 1994, 267(4): R901–R908. |
| [28] | 李振远, 李方方, 张勇, 等. 胃饥饿素:生物学特征及对动物采食量的调控[J]. 动物营养学报, 2016, 28(1) :35–42. |
| [29] | GNANAPAVAN S, KOLA B, BUSTIN S A, et al. The tissue distribution of the mRNA of ghrelin and subtypes of its receptor, GHS-R, in humans[J]. The Journal of Clinical Endocrinology & Metabolism, 2002, 87(6): 2988. |
| [30] | SMITH M W, TURVEY A, FREEMAN T C. Appearance of phloridzin-sensitive glucose transport is not controlled at mRNA level in rabbit jejunal enterocytes[J]. Experimental Physiology, 1992, 77(3): 525–528. DOI: 10.1113/expphysiol.1992.sp003616 |
| [31] | GARRIGA C, MORETO M, PLANAS J M. Hexose transport in the apical and basolateral membranes of enterocytes in chickens adapted to high and low NaCl intakes[J]. The Journal of Physiology, 1999, 514(1): 189–199. DOI: 10.1111/tjp.1999.514.issue-1 |
| [32] | GARRIGA C, PLANAS J M, MORETÓ M. Aldosterone mediates the changes in hexose transport induced by low sodium intake in chicken distal intestine[J]. The Journal of Physiology, 2001, 535(1): 197–205. DOI: 10.1111/tjp.2001.535.issue-1 |
| [33] | LAVERTY G, BJARNADÓTTIR S, ELBRØND V S, et al. Aldosterone suppresses expression of an avian colonic sodium-glucose cotransporter[J]. American Journal of Physiology:Regulatory, Integrative and Comparative Physiology, 2001, 281(4): R1041–R1050. |
| [34] | 王修启, 陈杰. 钠葡萄糖共转运载体 (SGLT1) 研究进展[J]. 中国畜牧兽医, 2003, 30(6) :24–26. |
| [35] | 杨秀平. 动物生理学[M]. 2版. 北京: 高等教育出版, 2002: 54-56. |
| [36] | SINGER T D, FINSTAD B, MCCORMICK S D, et al. Interactive effects of cortisol treatment and ambient seawater challenge on gill Na+, K+-ATPase and CFTR expression in two strains of Atlantic salmon smolts[J]. Aquaculture, 2003, 222(1/2/3/4): 15–28. |