




2. 河北北方学院农林科技学院, 张家口 075000
2. School of Agriculture and Forestry Science and Technology, Hebei North College, Zhangjiakou 075000, China
沙葱(Allium mongolicum Regel,AMR)属百合科多年生草本植物,主要分布于我国新疆、青海、甘肃、宁夏、陕西、内蒙古等地,是我国西北地区人民喜爱的优良佳肴。沙葱中富含蛋白质、脂肪、矿物质、微量元素、多糖、黄酮类化合物、大蒜素等多种营养物质和生物活性物质。据蒙药典记载,沙葱具有降血压、降血脂、开胃消食、健肾壮阳、治便秘等特殊功效[1]。沙葱因其含有较高的粗蛋白质和代谢能、较低的中性洗涤纤维,是一种优等牧草[2]。饲粮中添加沙葱、沙葱多糖和沙葱黄酮能改善肉羊生长性能和瘤胃内环境[3-6],并提高其抗氧化和免疫能力[7-9]。饲粮中添加沙葱和沙葱黄酮能调控肉羊体脂的脂肪酸组成和肌内脂肪(intramuscular fat,IMF)含量[3, 10-11]。
羊肉因其肉质细嫩、脂肪和胆固醇含量少备受人们喜欢[12]。养羊的模式从传统放牧饲养转变为舍饲或半舍饲集约化饲养,虽然从量上满足了人们对羊肉的需求,但是羊肉品质在下降,其中表现之一就是脂肪的过多沉积。羊肉特有的膻味影响人们对其接受度,研究表明羊肉的膻味主要来源于脂肪[12-13]。因此通过营养手段调控羊肉肌内脂肪沉积从而改善羊肉风味显得尤为必要。本课题组前期研究了饲粮中添加沙葱和沙葱黄酮可以改善背最长肌中肌内脂肪沉积和脂肪酸组成[6-10]。本试验在前期研究的基础上,以小尾寒羊为模型,研究基础饲粮中分别添加沙葱、沙葱多糖、沙葱提取多糖后的滤渣对肉羊生长性能的影响,并从肌内脂肪沉积、脂肪代谢相关的血清生化和激素指标及相关基因mRNA表达量角度探究沙葱、沙葱多糖、沙葱提取多糖后的滤渣对肉羊脂肪代谢的调节作用,以期为沙葱及其提取物作为天然饲料添加剂用于优质羊肉的生产提供科学依据。
1 材料与方法 1.1 试验材料沙葱粉由阿拉善盟浩海生物科技有限公司提供,主要有效成分为多糖[14]和黄酮类化合物[10];沙葱多糖由本实验室按照扈瑞平[14]的方法从沙葱粉中提取,它是由D-葡萄糖、L-鼠李糖、D-半乳糖构成的杂多糖,纯度为65%;滤渣为从沙葱粉中提取多糖后的滤渣,主要有效成分为黄酮类化合物。
1.2 试验设计选用40只体况良好、月龄相近、体重35 kg左右、健康的小尾寒羊公羊,随机分为4个组,每组10只羊。对照组饲喂基础饲粮,试验组分别在基础饲粮中添加0.1%沙葱多糖(多糖组)、10 g/(d·只)沙葱粉(沙葱组)、10 g/(d·只)沙葱提取多糖后的滤渣(滤渣组)。基础饲粮组成及营养水平见表 1。沙葱粉和沙葱多糖饲喂量分别按照卢媛[2]和包雨洪[5]试验结果确定,滤渣饲喂量按照沙葱粉饲喂量添加。沙葱粉和滤渣分别分早晚2次加入各组的基础饲粮中,每次5 g,与当次饲喂基础饲粮的1/2混合均匀并投料,当饲粮基本吃完时,饲喂剩余的饲粮。
|
|
表 1 基础饲粮组成及营养水平(干物质基础) Table 1 Composition and nutrient levels of the basal diet (DM basis) |
试验期共75 d,其中预试期15 d,正试期60 d,每天07:00和18:00各饲喂1次,自由采食,自由饮水,并记录投料量和剩料量。试验结束当天清晨空腹称重,并采集血液,静置20 min后,离心收集血清于1.5 mL EP管中,置于-20 ℃冰箱中保存备用。正试期结束时每组随机屠宰6只,采集背最长肌,一部分置于冻存管中,-80 ℃冰箱中保存,用于测定脂肪代谢相关基因mRNA表达量;一部分用锡箔纸包好置于-20 ℃冰箱中,用于测定肌内脂肪含量。
1.4 测定指标 1.4.1 生长性能正试期第1天和最后1天清晨对所有试验羊空腹称重,并记录初始体重和宰前体重,计算平均日采食量、平均日增重和料重比,计算公式如下:
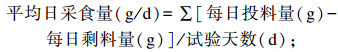
|
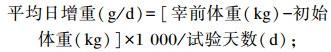
|

|
参照《食品中脂肪的测定》GB 5009.6—2016中的索氏抽提法测定。
1.4.3 血清脂肪代谢相关指标血清生化指标中总胆固醇(TC)、甘油三酯(TG)、高密度脂蛋白胆固醇(HDL-C)、低密度脂蛋白胆固醇(LDL-C)含量按照中生北控股份有限公司的试剂盒说明书采用全自动生化仪测定;血清激素指标中胰岛素(INS)、生长激素(GH)、瘦素(LEP)含量按照北京华英生物技术研究所的试剂盒说明书采用全自动放免计数仪测定。
1.4.4 背最长肌中脂肪代谢相关基因mRNA表达量采用实时荧光定量PCR测定背最长肌中与脂肪代谢相关的基因[激素敏感脂肪酶(HSL)、二酰甘油酰基转移酶1(DGAT1)、乙酰辅酶A羧化酶(ACC)、硬脂酰辅酶A去饱和酶(SCD)、脂肪酸合成酶(FAS)]mRNA表达量。根据NCBI中搜索的相应基因序列,利用Primer express 5.0进行引物设计,并进行特异性检测,由上海生工生物工程有限公司合成,基因引物序列信息见表 2,以甘油醛-3-磷酸脱氢酶(GAPDH)为管家基因。
|
|
表 2 基因引物序列信息 Table 2 Gene primer sequence information |
取50~100 mg样品放入经液氮冷却的研钵中,加入液氮研磨至粉末,用RNAiso plus(9109,TaKaRa)按照说明书提取RNA。按照反转录试剂盒(RR047A,TaKaRa)说明书将RNA反转录为cDNA,按照SYBR® Premix Ex TaqTM Ⅱ(RR820A,TaKaRa)说明书进行实时荧光定量PCR。扩增条件为预变性95 ℃,30 s;95 ℃,5 s,60 ℃,30 s,共40个循环。目的基因mRNA的表达量采用2-△△CT法计算。
1.5 数据分析采用SAS 9.0软件中的GLM程序进行方差分析,并用Duncan氏法进行多重比较,以P < 0.05作为差异显著性判断标准,0.05≤P < 0.10为有增加或降低的趋势,试验结果以平均值±标准差(mean±SD)表示。
2 结果 2.1 沙葱及其提取物对小尾寒羊生长性能的影响由表 3可知,与对照组相比,3个试验组的平均日采食量分别增加了4.88%、2.14%和7.09%,其中滤渣组显著增加(P < 0.05),多糖组和沙葱组均有增加趋势(0.05≤P < 0.10);3个试验组的平均日增重分别增加了13.65%、20.63%和25.46%,其中沙葱组和滤渣组均有增加趋势(0.05≤P < 0.10),多糖组差异不显著(P>0.05);3个试验组的料重比分别降低了3.11%、8.96%和8.95%,但均差异不显著(P>0.05)。
|
|
表 3 沙葱及其提取物对小尾寒羊生长性能的影响 Table 3 Effects of AMR and its extract on growth performance of small tail Han sheep |
由图 1可知,与对照组相比,3个试验组的背最长肌中肌内脂肪含量分别降低了8.22%、2.44%、5.67%,但均差异不显著(P>0.05)。

|
数据柱无字母标注表示差异不显著(P>0.05)。 Value columns with no letter mean no significant difference (P>0.05). 图 1 沙葱及其提取物对小尾寒羊背最长肌中肌内脂肪含量的影响 Fig. 1 Effects of AMR and its extract on longissimus dorsi intramuscular fat content of small tail Han sheep |
由表 4可知,与对照组相比,与脂肪代谢相关的血清生化指标中,3个试验组的血清TC、TG、HDL-C、LDL-C含量均没有显著差异(P>0.05),但是多糖组的血清TC和HDL-C含量有升高趋势(0.05≤P < 0.10);与脂肪代谢相关的血清激素指标中,3个试验组的血清LEP含量显著升高(P < 0.05),3个试验组的血清GH含量均有升高趋势(0.05≤P < 0.10),3个试验组的血清INS含量无显著差异(P>0.05)。3个试验组相比,多糖组的血清INS含量显著高于滤渣组(P < 0.05)。
|
|
表 4 沙葱及其提取物对小尾寒羊血液中脂肪代谢相关指标的影响 Table 4 Effects of AMR and its extract on serum lipid metabolism related indices of small tail Han sheep |
由表 5可知,与对照组相比,多糖组和滤渣组的DGAT1 mRNA表达量显著降低(P < 0.05),沙葱组和滤渣组的ACC、SCD mRNA表达量均显著降低(P < 0.05);沙葱组和滤渣组的FAS mRNA表达量有降低趋势(0.05≤P < 0.10)。与脂肪分解代谢相关基因HSL mRNA表达量与对照组相比,多糖组的HSL mRNA表达量显著升高(P < 0.05)。
|
|
表 5 沙葱及其提取物对小尾寒羊背最长肌中脂肪代谢相关基因mRNA表达量的影响 Table 5 Effects of AMR and its extract on mRNA expression levels of lipid metabolism related genes in longissimus dorsi of small tail Han sheep |
提高平均日增重和降低料重比是养殖业面临的主要问题,这将决定养殖业的经济效益。本试验结果表明,在基础饲粮中分别添加沙葱、沙葱多糖和沙葱提取多糖后的滤渣饲喂小尾寒羊时虽不能显著改善其生长性能,但不同程度增加了平均日增重和降低了料重比,这与赵国芬[3](基础饲粮中添加沙葱)、蔺婷娟[4](基础饲粮中添加沙葱多糖)、陈仁伟[10](基础饲粮中添加沙葱黄酮)的研究结果一致。机体免疫性能和抗氧化能力的提高,可以减少感染疾病的几率,使机体能充分利用营养物质,从而可能促进生长[15]。前期研究表明,沙葱、沙葱多糖和沙葱黄酮可提高肉羊抗氧化能力和免疫性能[7-9]。这说明饲粮中添加沙葱、沙葱多糖和沙葱提取多糖后的滤渣可不同程度增加平均日增重和降低料重比,可能与它们均能提高肉羊的免疫性能和抗氧化能力有关。
3.2 沙葱及其提取物对小尾寒羊脂肪代谢的调节作用羊肉特有的膻味影响人们对其接受度,研究表明羊肉的膻味主要来源于脂肪。本试验中3个试验组的背最长肌中的肌内脂肪含量虽没有显著变化,但有一定程度降低。这说明饲喂沙葱、沙葱多糖和沙葱提取多糖后的滤渣均可一定程度改善羊肉的膻味。卢媛[2]研究发现,在基础饲粮中分别添加10和15 g/(d·只)沙葱粉饲喂肉羊有降低肌内脂肪含量的趋势,而基础饲粮中添加20 g/(d·只)沙葱粉饲喂肉羊有升高肌内脂肪含量的趋势,这说明沙葱对肌内脂肪沉积量的影响受沙葱添加剂量的影响。
血清生化指标可以反映动物的生理代谢状态,并且与营养状况密切相关[16]。TC、TG、HDL-C、LDL-C是与脂肪代谢相关的血清生化指标[17]。本研究结果表明,3个试验组的血清TC、TG、HDL-C、LDL-C含量均没有显著变化,这说明沙葱、沙葱多糖和沙葱提取多糖后的滤渣对脂肪代谢相关的血清生化指标没有影响。LEP、GH和INS是与脂肪代谢相关的血清激素指标,这些激素可以影响机体中脂肪的代谢。INS可促进组织对葡萄糖与氨基酸的摄入,增加蛋白质与脂肪的合成,抑制蛋白质与脂肪的分解[18]。GH可促进物质代谢与机体生长,加速蛋白质合成,促进脂肪分解,使血清葡萄糖含量升高,同时促进肌肉细胞分裂增殖[16]。LEP是由脂肪细胞合成并分泌的具有多种功能的肽类激素,可以抑制食欲,增加能量消耗,抑制脂肪合成,促进脂肪分解,从而降低脂肪沉积[19-20]。本研究结果表明,3个试验组的血清LEP含量均显著升高,3个试验组的血清GH含量均有升高趋势,3个试验组的血清INS含量均无显著变化。这暗示3个试验组肌内脂肪变化可能与血清LEP含量降低有关。
基因是体内调节脂肪代谢的主要因素之一。DGAT1、ACC、SCD、FAS等是与脂肪合成代谢相关的基因。DGAT1基因编码二酰甘油酰基转移酶,二酰甘油酰基转移酶在三酯甘油的合成中起着重要作用[21]。FAS基因编码FAS,ACC基因编码ACC,ACC和FAS是参与脂肪酸从头合成的关键酶[22]。SCD基因编码SCD,SCD是合成单不饱和脂肪酸的限速酶[23]。HSL基因编码HSL,HSL负责脂肪的分解。植物多糖和黄酮类化合物可以通过调控脂肪合成和分解代谢相关基因的表达,从而影响脂肪代谢,但是这种调控受到植物多糖和黄酮类化合物剂量的影响[24-25]。本研究结果表明,与脂肪合成代谢相关基因中多糖组和滤渣组的DGAT1 mRNA表达量显著降低,沙葱组和滤渣组的ACC、SCD mRNA表达量均显著降低,沙葱组的DGAT1 mRNA表达量、多糖组的ACC和SCD mRNA表达量有一定程度降低,沙葱组和滤渣组的FAS mRNA表达量有降低趋势,多糖组的与脂肪分解代谢的相关基因HSL mRNA表达量显著升高,这说明沙葱、沙葱多糖、沙葱提取多糖后的滤渣可以通过下调脂肪合成相关基因和上调脂肪分解相关基因的表达共同调控脂肪代谢,但是它们对相关基因的调节作用有所不同,原因可能是它们所含沙葱多糖和黄酮类化合物含量不同导致的。有研究得到SCD基因是LEP信号的靶基因,LEP可抑制SCD基因表达[26]。本试验中3个试验组的SCD mRNA表达量和血清LEP含量都呈现下降趋势,这暗示沙葱、沙葱多糖和沙葱提取多糖后的滤渣可能是通过提高血清LEP含量来抑制SCD基因表达。
4 结论① 基础饲粮中分别添加沙葱、沙葱多糖、沙葱提取多糖后的滤渣饲喂小尾寒羊均不能显著改善其生长性能。
② 基础饲粮中分别添加沙葱、沙葱多糖、沙葱提取多糖后的滤渣饲喂小尾寒羊均不能显著改善背最长肌中肌内脂肪的沉积,但可以显著调节与脂肪代谢相关的血清LEP含量,并可以不同程度调节与脂肪代谢相关基因DGAT1、ACC、SCD、FAS和HSL的mRNA表达量。
| [1] |
SCHMITT B, SCHULZ H, STORSBERG J, et al. Chemical characterization of Allium ursinum L. depending on harvesting time[J]. Journal of Agricultural and Food Chemistry, 2005, 53(18): 7288-7294. DOI:10.1021/jf0504768 |
| [2] |
卢媛.沙葱、地椒风味活性成分及其对绵羊瘤胃发酵和羊肉风味的影响[D].硕士学位论文.呼和浩特: 内蒙古农业大学, 2002: 36-64. http://cdmd.cnki.com.cn/Article/CDMD-10129-2002090127.htm
|
| [3] |
赵国芬.沙葱和油料籽实对绵羊瘤胃发酵、营养物质消化流通及胴体品质影响的研究[D].博士学位论文.呼和浩特: 内蒙古农业大学, 2005: 92-151. http://cdmd.cnki.com.cn/Article/CDMD-10129-2006101290.htm
|
| [4] |
蔺婷娟.沙葱多糖对肉羊血液生化指标和肠道粘膜免疫影响的研究[D].硕士学位论文.呼和浩特: 内蒙古农业大学, 2010: 37-38. http://cdmd.cnki.com.cn/Article/CDMD-10129-1011178647.htm
|
| [5] |
包雨洪.沙葱多糖对绵羊瘤胃发酵、纤维素降解及血液生化指标的影响[D].硕士学位论文.呼和浩特: 内蒙古农业大学, 2009: 22-42. http://cdmd.cnki.com.cn/Article/CDMD-10129-2009131410.htm
|
| [6] |
包玲玲.沙葱黄酮类化合物对绵羊瘤胃发酵、内环境参数及纤维素降解的影响[D].硕士学位论文.呼和浩特: 内蒙古农业大学, 2015: 27-39. http://cdmd.cnki.com.cn/Article/CDMD-10129-1015427865.htm
|
| [7] |
宋巴达玛.沙葱多糖对肉羊免疫机能和抗氧化作用的影响[D].硕士学位论文.呼和浩特: 内蒙古农业大学, 2010: 42. http://cdmd.cnki.com.cn/Article/CDMD-10129-2010150485.htm
|
| [8] |
木其尔, 敖长金, 萨茹丽, 等. 沙葱总黄酮对肉羊抗氧化能力的影响[J]. 动物营养学报, 2016, 28(6): 1823-1831. DOI:10.3969/j.issn.1006-267x.2016.06.025 |
| [9] |
陈圣阳, 敖长金, 郑彦楷, 等. 沙葱黄酮对肉羊生产性能和肠道组织β-防御素基因表达的影响[J]. 动物营养学报, 2018, 30(3): 1095-1102. DOI:10.3969/j.issn.1006-267x.2018.03.034 |
| [10] |
陈仁伟.沙葱黄酮对肉羊生产性能及其肉品质的影响[D].硕士学位论文.呼和浩特: 内蒙古农业大学, 2016: 23-24. http://cdmd.cnki.com.cn/Article/CDMD-10129-1016249868.htm
|
| [11] |
MORÁN L, GIRÁLDEZ F J, PANSERI S, et al. Effect of dietary carnosic acid on the fatty acid profile and flavour stability of meat from fattening lambs[J]. Food Chemistry, 2013, 138(4): 2407-2414. DOI:10.1016/j.foodchem.2012.12.033 |
| [12] |
张利平, 吴建平. 肉羊体脂脂肪酸与肉品质关系的研究[J]. 甘肃农业大学学报, 2000, 35(4): 363-369. DOI:10.3969/j.issn.1003-4315.2000.04.001 |
| [13] |
WATKINS P J, KEARNEY G, ROSE G, et al. Effect of branched-chain fatty acids, 3-methylindole and 4-methylphenol on consumer sensory scores of grilled lamb meat[J]. Meat Science, 2014, 96(2): 1088-1094. DOI:10.1016/j.meatsci.2012.08.011 |
| [14] |
扈瑞平.沙葱多糖的分离、纯化和结构鉴定及其生物学活性的研究[D].博士学位论文.呼和浩特: 内蒙古农业大学, 2010: 27, 106-107. http://cdmd.cnki.com.cn/Article/CDMD-10129-2010147169.htm
|
| [15] |
杨耀翔, 杨玉, 董晓芳, 等. 苜蓿多糖对不同性别肉仔鸡生长性能、屠宰性能、肉品质及抗氧化性能的影响[J]. 动物营养学报, 2017, 29(2): 488-501. DOI:10.3969/j.issn.1006-267x.2017.02.016 |
| [16] |
赵天章.日粮油脂类型对羊肉脂肪酸和肌内脂肪含量的影响及其机理[D].博士学位论文.北京: 中国农业大学, 2014: 45-46. http://cdmd.cnki.com.cn/Article/CDMD-10019-1014221308.htm
|
| [17] |
韩丽, 潘杰, 张婷, 等. 低聚木糖对生长肥育猪血浆生化参数和肌肉脂肪酸组成的影响[J]. 动物营养学报, 2017, 29(9): 3316-3324. DOI:10.3969/j.issn.1006-267x.2017.09.036 |
| [18] |
MUOIO D M, DOHM G L, TAPSCOTT E B, et al. Leptin opposes insulin's effects on fatty acid partitioning in muscles isolated from obese ob/ob mice[J]. The American Journal of Physiology, 1999, 276(5): E913-E921. |
| [19] |
TRAYHURN P. Hypoxia and adipose tissue function and dysfunction in obesity[J]. Physiological Reviews, 2013, 93(1): 1-21. DOI:10.1152/physrev.00017.2012 |
| [20] |
GE J F, QI C C, ZHOU J N. Imbalance of leptin pathway and hypothalamus synaptic plasticity markers are associated with stress-induced depression in rats[J]. Behavioural Brain Research, 2013, 249: 38-43. DOI:10.1016/j.bbr.2013.04.020 |
| [21] |
刘文艳, 王长法, 仲跻峰, 等. 二脂酰甘油酰基转移酶1基因的研究进展[J]. 中国畜牧兽医, 2010, 37(7): 54-57. |
| [22] |
DERVISHI E, SERRANO C, JOY M, et al. The effect of feeding system in the expression of genes related with fat metabolism in semitendinous muscle in sheep[J]. Meat Science, 2011, 89(1): 91-97. DOI:10.1016/j.meatsci.2011.04.003 |
| [23] |
张蕊, 张宜辉, 邵丹, 等. 硬脂酰辅酶A去饱和酶基因的功能与调控[J]. 生命科学, 2013, 25(4): 378-382. |
| [24] |
朱晓娟.松针多糖和葛根黄酮组合对3T3-L1细胞分化及脂肪代谢的影响[D].硕士学位论文.南昌: 江西农业大学, 2016: 26-39. http://cdmd.cnki.com.cn/Article/CDMD-10410-1016280727.htm
|
| [25] |
刘姚.青钱柳多糖对3T3-L1脂肪细胞的细胞增殖及脂肪代谢的影响[D].硕士学位论文.南昌: 江西农业大学, 2013: 40-54.
|
| [26] |
KAKUMA T, LEE Y, HIGA M, et al. Leptin, troglitazone, and the expression of sterol regulatory element binding proteins in liver and pancreatic islets[J]. Proceedings of the National Academy of Sciences of the United States of America, 2000, 97(15): 8536-8541. DOI:10.1073/pnas.97.15.8536 |