




粗饲料是反刍动物饲粮的主要组分,通常占饲粮组成的40%~80%,其品质直接影响动物的健康与生产性能。饲料脂质是反刍动物的重要营养源,不仅是高能量物质,而且其脂肪酸组成影响反刍动物胴体、乳品质及健康[1-3]。因而,准确评价粗饲料的脂质组成与消化性具有重要的理论与实践意义。目前,我国和NRC(2001, 2016)[4-5]通常用粗脂肪(EE)表达脂质的含量,并视反刍动物对EE及其中的总脂肪酸(TFA)的消化利用率分别为95%和100%[4-7]。由于未能充分考虑脂质组成的复杂性及反刍动物特殊的消化道结构及消化生理,因而不能较好地反映饲料脂质的营养价值。
较新的康奈尔净碳水化合物-蛋白质体系(CNCPS 6.5)[8]引入Moate等[9]方法评价脂质营养价值。该法通过准确的化学分析方法对饲料脂质组分含量进行分析,利用瘤胃降解速率、脂肪酸生物氢化和合成速率,结合消化道流通速度模型等估计出不同脂肪酸的小肠消化吸收常数,预测饲料脂质的小肠可利用率,对饲料脂质的生物学价值做出有效、准确的预测。此法用于指导奶牛生产,取得了很好的效果。我国对该方法的应用研究不多。近年来,杨春英等[10]、冯德庆等[11]、张晓庆等[12]和那亚等[13]分析了油料籽实和牧草的脂肪酸组成,但尚未见对来源于纤维性农副产品的粗饲料脂质组成的分析。另外,因饲料脂质的消化吸收率不易直接测定,故这方面的报道也相对缺乏。因此,本研究旨在应用CNCPS 6.5提出的方法与原理,评定常用粗饲料脂质营养价值,以期为指导反刍动物生产提供基础数据。
1 材料与方法 1.1 样品采集与制备从河南省不同地区采集玉米秸秆、玉米叶、玉米茎、玉米秸黄贮、全株玉米青贮、花生蔓、大豆秧、红薯叶、红薯茎、西瓜秧10个样品,采集时尽量避免叶片脱落,保持原样的茎叶比例。新鲜饲料切成1 cm左右长度,混匀,按四分法取得次级样品。次级样品经120 ℃灭酶15 min后风干,粉碎,过1 mm筛,混匀,贮存于样品瓶已备分析。粗饲料的采样信息见表 1。
|
|
表 1 10种粗饲料的采样信息 Table 1 Sample information of 10 kinds of roughages |
样品干物质(DM)、EE含量测定分别按照GB/T 6435—2006、GB/T 6433—2006方法进行;脂肪酸含量测定按照GB/T 21514—2008方法进行。脂肪酸纯品的甲酯制备按照GB/T 17376—2008的方法进行。仪器:安捷伦6890A。色谱柱:BPX-70,毛细管分析柱(30.0 m×320 μm×0.25 μm)。气相色谱分析条件:进样口温度为210 ℃;柱箱温度为190 ℃;检测器温度为300 ℃;氮气流速为1.0 mL/min;氢气流速为30 mL/min;空气流速为400 mL/min。每个样品测定3次,取其平均值。
1.3 饲料脂质组分划分与消化吸收率计算应用CNCPS 6.5原理与方法[1, 3],将饲料脂质划分为脂肪酸、“甘油”和“色素”3个组分。其中,“甘油”定义为甘油三酯的醇部分和糖脂分子的糖残基集合,含量为TFA的0.11倍,瘤胃与瘤胃后的消化吸收率为100%;“色素”定义为色素、蜡质、烷类和固醇的集合体,计算为饲料EE扣除TFA和“甘油”后剩余的部分,假定其没有营养价值且完全以粪的形式排出;脂肪酸是营养上最重要的组分,假定饲料中脂肪酸以单、二或三甘油酯及糖脂或脂肪酸钙盐形式存在,不同脂肪酸具有各异的小肠消化吸收常数:辛酸(C8 : 0)、葵酸(C10 : 0)、月桂酸(C12 : 0)、豆蔻酸(C14 : 0)、棕榈酸(C16 : 0)、棕榈一烯酸(C16 : 1)、硬脂酸(C18 : 0)、顺式油酸(C18 : 1c)、反式油酸(C18 : 1t)、亚油酸(C18 : 2)、亚麻酸(C18 : 3)和20个碳原子及其以上的脂肪酸依次为1.000、1.000、0.954、0.751、0.725、0.640、0.728、0.893、0.786、0.830、0.775、0.587。
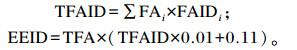
|
式中:TFAID与EEID分别为饲料TFA与EE小肠消化率(%);FAi为i脂肪酸占TFA的百分含量;FAIDi为i脂肪酸的小肠消化吸收系数;TFA为饲料脂质中TFA含量(%)。
1.4 数据处理与统计分析试验数据采用Excel 2003进行初步整理,用SPSS 20.0软件进行单因素方差分析(one-way ANOVA),用LSD模块进行多重比较,结果以平均值±标准差表示。P<0.05为差异显著。
2 结果与分析 2.1 10种粗饲料脂质含量与组成EE是表达饲料脂质营养价值的基本指标。由表 2可知,10种粗饲料EE含量变异范围为8.42~28.04 g/kg DM,大豆秧最高,显著高于其他粗饲料(P<0.05),玉米茎最低,显著低于玉米秸秆、全株玉米青贮、红薯叶、红薯茎(P<0.05)。5种玉米秸秆类饲料的EE含量变异范围为8.42~24.21 g/kg DM,玉米秸秆和全株玉米青贮显著高于其他3种(P<0.05),其他3种之间差异不显著(P>0.05)。花生蔓、西瓜秧、红薯叶、红薯茎和大豆秧5种粗饲料的EE含量依次升高,变异范围为10.08~28.04 g/kg DM,最低值与最高值均出现在豆科秸秆中,大豆秧显著高于其他4种(P<0.05),花生蔓显著低于除西瓜秧外的其他3种(P<0.05)。
|
|
表 2 10种粗饲料粗脂肪含量与组成 Table 2 Contents of EE and its components of 10 kinds of roughages |
脂质不同组分含量大致反映了饲料脂质营养价值的高低。脂质的能量价值主要取决于TFA含量。由表 2可知,10种粗饲料TFA含量总体不高,变异范围为1.84~15.39 g/kg DM,相当于157.32~636.09 g/kg EE。玉米秸秆最高,显著高于其他9种粗饲料(P<0.05),西瓜秧最低,显著低于除花生蔓以外的8种粗饲料(P<0.05)。5种玉米秸秆类饲料的TFA含量变异范围为2.85~15.39 g/kg DM,玉米秸秆显著高于其他4种(P<0.05),玉米叶显著低于玉米茎和玉米秸秆(P<0.05)。剩余的5种粗饲料TFA含量变异范围为1.84~7.41 g/kg DM,含量高低顺序与EE不同,由低到高依次为西瓜秧、花生蔓、大豆秧、红薯叶、红薯茎,西瓜秧显著低于除花生蔓以外的其他3种(P<0.05),红薯茎显著高于除红薯叶以外的其他3种粗饲料(P<0.05)。10种粗饲料中无能量价值的“色素”含量较高,变异范围为3.47~21.96 g/kg DM,大豆秧和玉米茎分别显著高于或低于其他粗饲料(P<0.05)。
2.2 10种粗饲料脂质中脂肪酸含量与组成脂肪酸组成不仅影响脂质的能量价值,而且影响反刍动物胴体、乳品质及健康。由表 3可知,10种粗饲料EE中的不同脂肪酸含量存在明显差异。玉米秸黄贮的C14 : 0、玉米茎的C18 : 0、玉米秸秆的C18 : 1和玉米叶的≥C20含量分别显著高于其他9种粗饲料对应的脂肪酸含量,玉米秸秆、玉米秸黄贮和全株玉米青贮的C16 : 0含量显著高于其他7种粗饲料(P<0.05)。由表 4可见,红薯茎和叶的中链脂肪酸(MCFA;C8~12)占TFA比例较高,分别为34.40%和18.00%,显著高于其他8种粗饲料(P<0.05)。红薯茎和叶的长链脂肪酸(LCFA;≥C14)占TFA比例显著低于其他8种粗饲料(P<0.05),其他8种粗饲料之间差异不显著(P>0.05)。
|
|
表 3 10种粗饲料的脂肪酸含量 Table 3 Fatty acid contents of 10 kinds of roughage (n=3) |
|
|
表 4 10种粗饲料脂肪酸组成 Table 4 Fatty acid composition of 10 kinds of roughages (n=3) |
亚油酸与亚麻酸是必需脂肪酸。由表 3可知,10种粗饲料亚油酸含量变异范围为21.61~287.34 g/kg EE,相当于占TFA的9.08%~45.17%(表 4),玉米秸秆和玉米秸黄贮分别显著高于和低于其他9种粗饲料(P<0.05),玉米茎显著高于玉米叶(P<0.05),为玉米叶的1.59倍,但红薯茎、叶之间差异不显著(P>0.05)。10种粗饲料亚麻酸占TFA的变异范围为1.81%~31.39%,大豆秧显著高于其他9种粗饲料(P<0.05),玉米秸秆和玉米秸黄贮显著低于其他8种粗饲料(P<0.05),但玉米秸秆和玉米秸黄贮之间差异不显著(P>0.05),玉米与红薯叶亚麻酸含量分别是对应茎的2.05倍和1.99倍,差异显著(P<0.05)。
2.3 10种粗饲料脂质与脂肪酸小肠消化率表 5列出了基于CNCPS 6.5方法评价的EE与TFA小肠消化率。由表 5可知,10种粗饲料的EE小肠消化率变异范围为13.92%~59.64%,玉米秸秆最高,显著高于其他9种粗饲料(P<0.05),西瓜秧和大豆秧较低,显著低于其他8种粗饲料(P<0.05),最高和最低的粗饲料EE小肠消化率相差4.28倍。5种玉米秸秆类饲料的EE小肠消化率变异范围为26.22%~59.64%,除玉米叶和玉米秸黄贮之间差异不显著(P>0.05)外,其他粗饲料之间差异均显著(P<0.05)。剩余的5种粗饲料EE小肠消化率变异范围为13.92%~30.69%,除大豆秧与西瓜秧、红薯叶与茎之间差异不显著(P>0.05)外,其他粗饲料之间差异均显著(P<0.05)。
|
|
表 5 基于CNCPS方法评价的10种粗饲料EE与TFA小肠消化率 Table 5 EE and TFA small intestine digestibility of 10 kinds of roughages evaluated by CNCPS method (n=3) |
10种粗饲料的TFA小肠消化率变异范围为76.85%~85.15%,红薯茎最高,显著高于其他9种粗饲料(P<0.05)。5种玉米秸秆类饲料的TFA小肠消化率变异范围为76.85%~82.76%,玉米秸秆显著高于其他4种(P<0.05),玉米叶和玉米秸黄贮之间无显著差异(P>0.05)。剩余的5种粗饲料TFA小肠消化率变异范围为77.53%~85.15%,红薯茎显著高于其他4种粗饲料(P<0.05)。
3 讨论 3.1 10种粗饲料脂质含量与组成目前,国内有关粗饲料脂质组成研究不多。本研究发现,10种粗饲料TFA和色素含量变异范围分别为1.84~15.39 g/kg DM和3.47~21.96 g/kg DM,分别相当于占EE的15.73%~63.61%和29.39%~82.54%。10种粗饲料TFA含量不仅远低于动物油脂和油料籽实EE中TFA的含量(一般占88%)[14],而且不同原料间差异较大,最高与最低相差4.04倍。这一结果提示,NRC(2016)[5]将饲料EE消化率视为95%,可能仅适用于来源于动物油脂和油料籽实的EE,而不适用来源于纤维性农副产品的粗饲料中EE。本研究也发现,即使同为完熟期的玉米秸秆、玉米茎、玉米叶与玉米秸黄贮,其TFA含量变异范围为2.85~15.39 g/kg DM,变异也较大。随着收割时间的推迟,玉米秸秆中DM含量升高,TFA含量明显降低,这可能与收获迟的秸秆自身呼吸消耗造成了营养损失有关[15]。成熟期玉米秸秆与乳熟期全株玉米青贮TFA含量的差异,除生育期不同外,也可能与玉米秸秆和玉米穗中的TFA分布不均衡有关[16]。NRC(2001, 2016)[4-5]和CNCPS 6.5数据库均没有考虑不同生育期与加工方法的玉米秸秆TFA含量的差异,本研究为实际应用提供了数据参考。
3.2 10种粗饲料脂质中脂肪酸组成饲料脂质中的脂肪酸不仅是主要的供能物质,而且其中的MCFA具有快速供能、不易在机体沉积、抑制病原菌存活等特殊的生理效应[3]。食用植物油除椰子油外,其他几乎不含MCFA[10]。一般牧草与青贮饲料中也几乎不含MCFA[11-13, 17]。冯德庆等[11]发现红薯叶脂肪酸中含有7.7%的月桂酸,其他MCFA未测定。本研究发现,红薯茎、叶脂肪酸中分别含34.40%与18.00%的MCFA。这一结果提示,适量选用红薯秧作为反刍动物饲料原料,可能有助于生产出健康、安全的畜产品。
另外,共轭亚油酸(CLA)是一组在碳9、11或10、12位具有双键的亚油酸位置异构体和空间异构体的总称。有研究表明,9c,11t-CLA和10t,12c-CLA具有抗癌、抗氧化、降低胆固醇、免疫调节、抑制脂肪沉积、抗动脉粥样、促进成骨、降糖等多种特殊的生物学功能[18-19]。天然CLA主要来源于反刍动物中的含脂类物质,是膳食CLA的主要来源[20]。反刍动物64%以上的CLA由内源合成,饲料亚油酸与亚麻酸是合成的主要前体,并以后者作用更大[17, 21]。虽然一些油料籽实和植物油含有较高含量的亚油酸和亚麻酸[14],但缺点是成本较高,而且对瘤胃发酵及乳脂率有负面影响。相对油料籽实和植物油,植物茎叶来源广泛,成本低廉。由于植物茎叶亚油酸的双键位于碳7, 9位置,对于合成9c,11t-CLA的效率很低[21-22],植物茎叶亚麻酸是合成CLA的主要底物来源。本研究发现,一些来源于纤维性农副产品的粗饲料,如大豆秧、西瓜秧和红薯叶中含有较高含量的亚麻酸。其他研究[11-13, 17]也表明,多数饲草脂肪酸中含有较高含量的亚麻酸。因此,高亚麻酸含量粗饲料搭配适量的含亚油酸丰富的植物油,可能是提高反刍动物CLA含量的一个重要生产模式。
3.3 10种粗饲料脂质与脂肪酸小肠消化率因饲料脂质和脂肪酸消化率不易测定,有关研究报道不多。本研究发现,基于CNCPS方法评价的EE和TFA小肠消化率,来源于纤维性农副产品的粗饲料TFA小肠消化率变异范围为76.85%~85.15%,低于NRC(2001)[4]采用的100%。这主要是因为本研究考虑了反刍动物小肠对不同脂肪酸消化率差异。另外,测试粗饲料EE小肠消化率变异范围仅为13.92%~59.64%,远低于NRC(2016)[5]采用的95%。造成这种差异的原因主要是由于测定粗饲料EE中不可消化利用的“色素”含量高达41.31%~82.56%,其次是由于不同粗饲料脂肪酸组成不同引起的TFA消化率的差异。由于CNCPS 6.5评价饲料脂质营养价值不仅考虑了脂质组成,而且也考虑了不同脂肪酸消化率的差异,能更准确地评定反刍动物对不同饲料EE与TFA消化吸收率。建议加大该法在我国推广应用,以更精确地评价饲料脂质营养价值。
4 结论① 粗饲料EE和TFA含量较低,变异范围分别为8.42~28.04 g/kg DM和1.84~15.39 g/kg DM;不具能量价值的“色素”含量较高,变异范围为3.47~21.96 g/kg DM。
② 应用CNCPS方法评价粗饲料EE小肠消化率较低,变异范围为13.92%~59.64%;TFA小肠消化率较高,变异范围为76.85%~85.15%。
| [1] |
李秋凤, 许蕾蕾, 李建国, 等. 亚麻籽对育肥牛肉品质及脂肪酸的影响[J]. 草业学报, 2013, 22(5): 272-279. |
| [2] |
卜登攀, 王家启, DHIMAN T R, 等. 植物油来源亚油酸和亚麻酸对乳脂CLA合成的影响[J]. 畜牧兽医学报, 2007, 38(7): 663-671. DOI:10.3321/j.issn:0366-6964.2007.07.008 |
| [3] |
王建军, 王恬. 中链脂肪酸的生物学特性及其在动物生产中的应用[J]. 动物营养学报, 2011, 23(7): 1073-1078. DOI:10.3969/j.issn.1006-267x.2011.07.001 |
| [4] |
NRC.Nutrient requirements of dairy cattle[S]. 7th ed.Washington, D.C.: National Academy Press, 2001.
|
| [5] |
NRC.Nutrient requirements of beef cattle[S]. 8th ed.Washington, D.C.: National Academy Press, 2016.
|
| [6] |
SNIFFEN C J, O'CONNOR J D, VAN SOEST P J, et al. A net carbohydrate and protein system for evaluating cattle diets:Ⅱ.Carbohydrate and protein availability[J]. Journal of Animal Science, 1992, 70(11): 3562-3577. DOI:10.2527/1992.70113562x |
| [7] |
BOSTON R C, FOX D G, SNIFFEN C J, et al.The conversion of a scientific model describing dairy cow nutrition and production to an industry tool: the CPM dairy project[M]//MCNAMARA J, FRANCE J, BEEVER D.Modelling nutrient utilization in farm animals.Oxford: CABI Publishing, 2000: 361-377.
|
| [8] |
VAN AMBURGH M E, COLLAO-SAENZ E A, HIGGS R J, et al. The cornell net carbohydrate and protein system:updates to the model and evaluation of version 6.5[J]. Journal of Dairy Science, 2015, 98(9): 6361-6380. DOI:10.3168/jds.2015-9378 |
| [9] |
MOATE P J, CHALUPA W, JENKINS T C, et al. A model to describe ruminal metabolism and intestinal absorption of long chain fatty acids[J]. Animal Feed Science and Technology, 2004, 112(1/2/3/4): 79-105. |
| [10] |
杨春英, 刘学铭, 陈智毅. 15种食用植物油脂肪酸的气相色谱-质谱分析[J]. 食品科学, 2013, 34(6): 211-214. |
| [11] |
冯德庆, 黄勤楼, 李春燕, 等. 28种牧草的脂肪酸组成分析研究[J]. 草业学报, 2011, 20(6): 214-218. |
| [12] |
张晓庆, 张英俊, 闫伟红, 等. 克氏针茅草原七种植物脂肪酸组分及其变化[J]. 中国草地学报, 2013, 35(1): 116-120. DOI:10.3969/j.issn.1673-5021.2013.01.019 |
| [13] |
那亚, 孙启忠, 王红梅. 呼伦贝尔草甸草原牧草青贮饲料脂肪酸成分研究[J]. 草业学报, 2017, 26(2): 215-223. |
| [14] |
中国饲料数据库. 中国饲料成分及营养价值表(2017年第28版)(续)[J]. 中国饲料, 2017(22): 31-36. |
| [15] |
崔卫东, 董朝霞, 张建国, 等. 不同收割时间对甜玉米秸秆的营养价值和青贮发酵品质的影响[J]. 草业学报, 2011, 20(6): 208-213. |
| [16] |
张亚龙. 收获期对寒地青贮玉米营养价值的影响[J]. 玉米科学, 2007, 15(4): 123-124, 132. DOI:10.3969/j.issn.1005-0906.2007.04.032 |
| [17] |
李志强, 刘凤珍, 卢鹏, 等. 几种重要饲草的脂肪酸成分分析[J]. 中国奶牛, 2006(10): 3-6. DOI:10.3969/j.issn.1004-4264.2006.10.002 |
| [18] |
李凯, 周宁, 李郝宇, 等. 共轭亚油酸生理功能研究进展[J]. 食品研究与开发, 2012, 33(7): 226-228. DOI:10.3969/j.issn.1005-6521.2012.07.064 |
| [19] |
田玉霞, 苗榕宸, 杨丹白, 等. 共轭亚油酸甘油酯的研究进展[J]. 中国油脂, 2015, 40(6): 32-35. DOI:10.3969/j.issn.1003-7969.2015.06.009 |
| [20] |
成亮, 刘成国, 刘婷婷, 等. 共轭亚油酸的生理功能及提高其在乳制品中含量的途径[J]. 食品与机械, 2010, 26(3): 170-173. |
| [21] |
GRⅡNARI J M, CORL B A, LACY S H, et al. Conjugated linoleic acid is synthesized endogenously in lactating dairy cows by △9-desaturase[J]. The Journal of Nutrition, 2000, 130(9): 2285-2291. DOI:10.1093/jn/130.9.2285 |
| [22] |
魏宏阳, 王加启. 牛乳脂共轭亚油酸含量调控研究进展[J]. 中国饲料, 2003(7): 12-13, 17. DOI:10.3969/j.issn.1004-3314.2003.07.008 |