




木薯是一种直立灌木,广泛种植于热带地区,是当地的主要粮食和能源植物[1]。据报道,2015年我国木薯消费总量为939万t,预计到2020年,全世界木薯消费量将达到2.71亿t[2]。木薯的加工与利用伴随着木薯渣等副产物的大量产生,未及时处理的木薯渣会腐败恶化、污染环境并危害人和动物健康[3]。有研究表明,木薯渣中无氮浸出物含量高达60%,总能为16.37~17.75 MJ/kg[3],并含有丰富的氨基酸和铜、锌、锰、铁等矿物质[4]。此外,木薯渣与压片玉米等常规饲料原料相比具有价格低廉的优势,是一种有待进一步开发和利用的畜禽饲料原料。
包淋斌等[5]发现锦江黄牛瘤胃对木薯渣的干物质(DM)、有机物(OM)、中性洗涤纤维(NDF)、酸性洗涤纤维(ADF)和粗蛋白质(CP)有效降解率高于苔草和苜蓿,表明其在锦江黄牛中有较高的饲喂价值。Dos Santos Filho等[6]用木薯渣替代绵羊饲粮中的压片玉米,发现当替代比例为25%时,能够有效提高绵羊平均日增重;吕小康等[7]也研究发现,饲粮中添加木薯渣(低于20%)能够线性提高湖羊羔羊的生长性能,但会影响其抗氧化和肝脏功能。以上研究表明,木薯渣在肉牛和羊上具有替代苜蓿或压片玉米等饲料原料、提高动物生长性能的潜力,然而木薯渣在奶牛上的应用还有待进一步探索。泌乳中后期奶牛应适当控制对压片玉米等精料的摄入,以控制其体况[8],而木薯渣纤维含量高,可作为粗饲料,同时淀粉含量也高,亦可作为能量饲料替代玉米应用于奶牛饲粮中[4]。因此,本试验旨在通过体外瘤胃微生物发酵试验来探究木薯渣替代压片玉米对泌乳中后期奶牛的饲喂效果,为木薯渣适量替代压片玉米用作该生理阶段奶牛的饲料原料提供依据。
1 材料与方法 1.1 试验材料试验用木薯渣(产自泰国,提取淀粉后经物理压榨而成)由北京九正生物科技有限公司提供,压片玉米由北京中地畜牧科技有限公司提供。饲料样品在烘箱中65 ℃烘干48 h至恒重后在空气中回潮粉碎过1 mm分析筛,用于常规营养成分含量的测定及体外瘤胃微生物发酵试验,试验样品的常规营养成分含量如表 1所示。
|
|
表 1 木薯渣及压片玉米常规营养成分含量(风干基础) Table 1 Conventional nutrient contents of cassava residue and tablet corn (air-dry basis) |
本试验共设计7个组:分别用木薯渣以0(对照组)、5%、10%、15%、20%、25%和30%(试验组)的比例替代压片玉米(风干基础),每组4个重复,各组发酵底物原料组成及其营养水平见表 2。
|
|
表 2 各组发酵底物原料组成及营养水平(风干基础) Table 2 Composition and nutrient levels of fermentation substrate in each group (air-dry basis) |
本试验采用中国农业大学杨红建等[9]自主研发设计的AGRS-Ⅲ型微生物发酵产气系统,对累积产气量(GP)进行实时监测和记录。根据Menke等[10]提出的方法配制缓冲液,接种前缓慢持续通入二氧化碳(CO2)气体30 min左右,调节混合液pH至6.8,置于39 ℃恒温水浴锅备用。瘤胃液于试验当天晨饲前采自3头处于泌乳中后期、装有永久瘤胃瘘管的成年荷斯坦奶牛。试验牛的饲粮由CMP软件制作配方配制,其总净能为175.73 MJ/kg,泌乳净能7.32 MJ/kg,粗蛋白质含量为16.9%,试验牛干物质采食量为24.0 kg/d,每天饲喂2次,自由饮水。采集到的瘤胃液经4层纱布过滤后置于39 ℃恒温水浴锅,不断搅拌并持续通入CO2备用。称取0.5 g样品并准确移取50 mL缓冲液和25 mL瘤胃液于发酵瓶,通入CO2 8~10 s后,加上几丁质胶塞,上紧螺盖,与产气系统对应编号的通道气路接口连接,置于39 ℃恒温培养。分别于发酵3、6、12、24和48 h后关闭系统,依次取出发酵瓶测定发酵液pH,并将瓶中发酵物全部转入已烘干称重的尼龙袋(300目,9 cm×14 cm)中过滤,尼龙袋用水反复清洗至澄清后放入烘箱中65 ℃烘干至恒重后在空气中回潮称重,用于干物质消失率(IVDMD)的计算,滤液用于发酵液氨态氮(NH3-N)、微生物蛋白(MCP)及挥发性脂肪酸(VFA)含量的测定。
1.3 检测指标及方法 1.3.1 IVDMD、GP及产气动力学参数IVDMD根据发酵前后底物的干物质含量,利用差减法计算得到;总产气量由AGRS-Ⅲ装置自动记录GP并参照Ørskov等[11]提出的指数函数模型拟合得到:
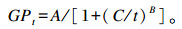
|
式中:GPt为t时间的总产气量(mL/g DM);A为发酵底物在该产气速率下的理论最大产气量(mL/g DM);B为底物发酵产气过程中的曲线拐点参数;C为产气量达到总产气量1/2时所用的时间(h);t为体外培养时间(h)。
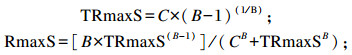
|
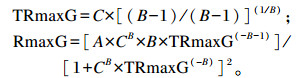
|
式中:TRmaxS为达到底物最大降解率所需的时间(h);RmaxS为底物的最大降解率(h-1);TRmaxG为达到最大产气速率所需的时间(h);RmaxG为最大产气速率(mL/h);A为发酵底物在该产气速率下的理论最大产气量(mL/g DM);B为底物发酵产气过程中的曲线拐点参数;C为产气量达到总产气量1/2时所用的时间(h)。
1.3.2 发酵液pH,NH3-N、MCP和VFA含量的测定发酵液pH的测定用梅特勒FiveEasy Plus系列pH计进行;NH3-N含量的测定根据Verdouw等[12]提出的亮蓝比色法进行,每个样品设3个平行;MCP含量的测定根据Makkar等[13]提出的考马斯亮蓝比色法进行,每个样品设3个平行;VFA含量的测定根据Erwin等[14]提出的气相色谱分析法进行,每个样品设2个平行。
1.4 数据统计分析试验所得数据用Excel 2007简单汇总整理后用SAS 9.2统计软件的NLIN程序拟合得到产气动力学参数A、B、C、TRmaxG、RmaxG、TRmaxS、RmaxS,并采用SAS 9.2 ANOVA程序进行单因素方差分析,用Duncan氏法进行多重分析,P≤0.05为差异显著。
2 结果与分析 2.1 不同比例木薯渣替代压片玉米对泌乳中后期奶牛体外瘤胃发酵48 h后GP、IVDMD及产气动力学参数的影响由表 3可知,随木薯渣替代压片玉米比例的增加,各组体外培养48 h后GP、理论最大产气量、最大产气速率均呈显著线性增加(P < 0.05),且30%组的最大产气速率显著高于0、5%、15%和25%组(P < 0.05)。此外,体外培养48 h后的IVDMD、曲线拐点参数、达到总产气量1/2所用的时间、底物最大的降解率所需的时间、底物最大降解率、达到最大产气速率所需的时间各组之间均无显著差异(P>0.05)。
|
|
表 3 不同比例木薯渣替代压片玉米对泌乳中后期奶牛体外瘤胃发酵产气量及动力学参数的影响 Table 3 Effects of different proportions of cassava residue substituted for tablet corn on in vitro rumen fermentation gas production and kinetic parameters of dairy cows in middle and late lactation |
由表 4可知,随发酵时间的延长,各组NH3-N含量呈现先下降后上升的趋势,并在体外培养12 h时各组NH3-N含量达到最低。体外培养24 h时,各组的NH3-N含量随木薯渣替代比例的增加呈显著线性增加(P < 0.05),且体外培养12 h时,0、25%、30%组的NH3-N含量显著高于5%、10%组(P < 0.05),与15%、20%组相比无显著差异(P>0.05);体外培养24 h时,30%组的NH3-N含量显著高于0、5%、10%、15%组(P < 0.05),与20%、25%组相比无显著差异(P>0.05)。
|
|
表 4 不同比例木薯渣替代压片玉米对泌乳中后期奶牛体外瘤胃发酵NH3-N含量的影响 Table 4 Effects of different proportions of cassava residue substituted for tablet corn on in vitro rumen fermentation NH3-N content of dairy cows in middle and late lactation |
由表 5可知,体外培养3 h时,各组的MCP含量随木薯渣替代比例的增加呈显著二次增加(P < 0.05),在其余各培养时间点,各组的MCP含量之间均无显著差异(P>0.05)。
|
|
表 5 不同比例木薯渣替代粉碎玉米对体外瘤胃发酵MCP含量的影响 Table 5 Effects of different proportions of cassava residue substituted for tablet corn on in vitro rumen fermentation MCP content of dairy cows in middle and late lactation |
由表 6可知,随发酵时间的延长,发酵液的pH整体呈下降趋势,其变化范围是6.25~7.68。体外培养24 h时,各组的pH随木薯渣替代比例的增加呈线性增加(P=0.05),在其余各培养时间点,各组的pH之间均无显著差异(P>0.05)。
|
|
表 6 不同比例木薯渣替代压片玉米对泌乳中后期奶牛体外瘤胃发酵液pH的影响 Table 6 Effects of different proportions of cassava residue substituted for tablet corn on in vitro rumen pH of dairy cows in middle and late lactation |
由表 7可知,各组丁酸、戊酸、总VFA含量随木薯渣替代比例的增加呈显著二次增加(P < 0.05),其余各挥发酸含量各组之间均无显著差异(P>0.05)。其中,丁酸含量20%组显著高于0、25%、30%组(P=0.05),与5%、10%、15%组相比无显著差异(P>0.05);戊酸、总VFA含量20%组均显著高于0、15%、25%、30%组(P < 0.05),与5%、10%组相比无显著差异(P>0.05)。
|
|
表 7 不同比例木薯渣替代压片玉米对泌乳中后期奶牛体外瘤胃发酵48 h VFA含量的影响 Table 7 Effects of different proportions of cassava residue substituted for tablet corn on in vitro rumen fermentation VFA content after cultivating for 48 h of dairy cows in middle and late lactation |
干物质降解率是影响奶牛干物质采食量的重要因素,它可表示反刍动物摄入饲粮中有机物的可利用程度,也是衡量饲粮营养价值的重要指标。在一定时间内,奶牛的干物质降解率越高,其干物质采食量就越大[15-16]。本试验中,不同比例木薯渣替代压片玉米后,IVDMD数值上有所增加,这可能与木薯渣替代比例的增加为瘤胃微生物的发酵提供了更为适宜的底物碳氮比例,从而微生物对发酵底物的利用率提高有关[17]。此外,饲粮在发酵过程中,瘤胃微生物会消耗动物摄入的碳水化合物及其他营养物质从而产生甲烷、氢气、CO2等气体,一定时间内的GP是反映底物被瘤胃微生物利用程度和底物营养价值高低的重要指标[18]。体外发酵产气量与底物所含碳水化合物的降解速率密切相关,产气量越大则表明底物发酵程度越好、瘤胃消化程度越高。有研究表明,体外发酵产气量与发酵底物中的NDF含量呈负相关[18],而木薯渣中含有大量不易发酵的NDF和ADF,但本试验中体外培养48 h后的GP随木薯渣代替比例的增加呈线性增加,这可能是因为2种饲料原料混合后,易发酵的营养成分和不易发酵的结构性碳水化合物的比例较单一饲料更为合理,使得微生物的活性更强,因而更易被瘤胃微生物利用[17]。因此,在后续研究中,可以进一步关注木薯渣替代压片玉米对泌乳中后期奶牛的瘤胃微生物菌群组成和微生物活性的影响。
3.2 不同比例木薯渣替代压片玉米对泌乳中后期奶牛体外瘤胃发酵NH3-N、MCP含量的影响瘤胃液中的NH3-N是瘤胃氮代谢过程的重要产物,也是合成MCP的主要氮源,NH3-N含量变化是衡量瘤胃微生物对饲粮中氮的利用和MCP合成是否平衡的重要指标[15]。有研究表发现,瘤胃液中的NH3-N含量与底物中粗蛋白质的含量呈正相关,且底物中的淀粉含量越高,瘤胃微生物对其中蛋白质的降解率越低[19]。本试验中,在体外培养24 h时,NH3-N含量随木薯渣替代比例的呈线性增加现象,且体外培养24 h时,30%组的NH3-N含量显著高于0、5%、10%、15%组,这可能是与木薯渣的淀粉含量低于压片玉米,故随木薯渣替代比例的增加,发酵底物中的淀粉含量随之降低,促进瘤胃微生物降解蛋白质产生NH3-N有关。还可能与随着木薯渣替代比例的增加,发酵底物的能氮比例更趋于平衡、能促进瘤胃微生物的生长、加速底物含氮物质的分解有关,这与卜登攀等[20]提出的“瘤胃微生物对含氮物质和能量载体具有依赖性、选择性和时效性,是能氮同步性结果的最直接指标”的观点一致。
MCP为反刍动物提供了60%~70%的蛋白质需要量,是反刍动物最主要的氮源,也是表征瘤胃微生物对底物中氮的利用效率和瘤胃微生物菌群数量的重要指标[21]。本试验中,在除体外培养12 h外的各时间点,均是30%组的MCP含量最大,且体外培养24 h时,随木薯渣替代比例的增加,MCP含量呈增加趋势,这与此时NH3-N含量随木薯渣替代比例增加线性增加、为MCP的合成提供了充足的氮源密切相关。还有研究发现,MCP的合成主要取决于瘤胃内碳水化合物和蛋白质的利用效率,当瘤胃中氨和能量的释放不同步时,可发酵底物的利用率会有所下降,从而导致MCP的合成量降低[22-23]。此外,MCP的合成同时需要能量和碳架[24],碳水化合物和粗纤维均可为MCP的合成提供碳架[25],且只有当氨的释放速度与能量和碳架的释放速度同步时,微生物的固氮作用最佳[17]。因此,本试验中MCP含量随木薯渣替代比例的增加而增加的现象可能还与随着木薯渣替代比例的增加,改善了底物中氮、能量和碳架释放的同步性有关。
3.3 不同比例木薯渣替代压片玉米对泌乳中后期奶牛体外瘤胃发酵液pH、VFA含量的影响瘤胃液的pH是反映瘤胃发酵水平的一项综合指标,它受饲粮性质、唾液分泌、渗透压、瘤胃内VFA及其他有机酸的含量、瘤胃中的水流量和食糜流速以及饲粮缓冲力等诸多因素的影响。奶牛借助复杂的酸碱平衡/调节系统,将瘤胃pH维持在5.5~7.5[26]。本试验在体外培养条件下,用不同比例的木薯渣替代压片玉米,随时间的延长,由于发酵产生的VFA的不断积累,各组在各时间点的发酵液pH逐渐降低,但均在正常范围内,即可说明用木薯渣替代压片玉米能够为泌乳中后期奶牛的瘤胃微生物提供稳定、适宜的生长环境。
VFA是瘤胃微生物发酵的重要产物,能为反刍动物提供70%~80%的能量[27]。其中乙酸是反刍动物脂肪合成的主要前体物质,是乳腺合成乳脂中短链脂肪酸的重要原料,乙酸转化为乳脂的效率为67%~71%[28],而丙酸是葡萄糖合成的重要前体物质,主要用于体脂和乳糖的合成,丙酸的生成可以竞争性地消耗氢气,从而有效抑制甲烷的合成[29]。乙酸/丙酸可以反映反刍动物对能量的利用情况和饲料发酵模式,其比例应大于2.2 : 1.0[15],其中丙酸的产生速率与饲料氮的沉积呈正比,乙酸与丙酸产量的不匹配会影响氮的沉积[30]。本试验中体外培养48 h后,20%组的乙酸和丙酸含量数值上均为各组最高,据此可推断在此替代比例下,木薯渣对奶牛瘤胃发酵类型无不良影响,并且还能够促进乳脂的合成、竞争性地消耗氢气,降低甲烷的生成,提高饲料氮的沉积效率,减少粪氮和尿氮的排出量。此外,各组丁酸、戊酸、总VFA含量随木薯渣替代比例的增加呈二次增加,且20%组显著高于25%、30%组,说明了该比例下非结构性碳水化合物的发酵率高,可生成大量的VFA,为微生物合成MCP提供能量。还有研究表明,戊酸、异丁酸和异戊酸分别是饲料蛋白质中缬氨酸、亮氨酸和异亮氨酸的微生物发酵产物[31],本试验中20%组的这3种VFA含量均为各组最高,即可在一定程度上说明该在木薯渣替代比例为20%时,可以促进瘤胃微生物对底物中这3种氨基酸的代谢,而且异丁酸和异戊酸属于支链VFA,可以刺激粗纤维分解菌的活性,增大结构性碳水化合物分解菌的生物量,提高干物质的消化率[32]。
4 结论本试验条件下,木薯渣替代压片玉米的比例不高于30%时对泌乳中后期奶牛体外瘤胃发酵特性无不良影响,且在替代比例为20%时可显著提高发酵液丁酸、戊酸和总VFA含量,在替代比例为30%时可显著提高发酵24 h时的NH3-N含量和48 h时的GP。因此,由体外试验可知,木薯渣替代压片玉米的比例为20%~30%较为适宜,但此试验结果仍需通过奶牛体内试验进一步验证。
| [1] |
马思雅, 顾进宝, 马晓雯, 等. 木薯SR家族基因的克隆和亚细胞定位[J]. 分子植物育种, 2018, 16(9): 2768-2777. |
| [2] |
张芹, 李广利, 于迎辉, 等. 我国木薯深加工现状及发展分析[J]. 粮食与饲料工业, 2017, 12(1): 31-34. |
| [3] |
冀凤杰, 王定发, 侯冠彧, 等. 木薯渣饲用价值分析[J]. 中国饲料, 2016(6): 37-40. |
| [4] |
崔艺燕, 田志梅, 李贞明, 等. 木薯及其副产品的营养价值及在动物生产中的应用[J]. 中国畜牧兽医, 2018, 45(8): 2135-2146. |
| [5] |
包淋斌, 黎力之, 潘柯, 等. 木薯渣在锦江黄牛瘤胃中的降解率测定[J]. 中国饲料, 2015(21): 12-15. |
| [6] |
DOS SANTOS FILHO H, VÉRAS R M, DE ANDRADE FERREIRA M A, et al. Liquid residue of cassava as a replacement for corn in the diets of sheep[J]. Tropical Animal Health and Production, 2015, 47(6): 1083-1088. DOI:10.1007/s11250-015-0830-7 |
| [7] |
吕小康, 王杰, 王世琴, 等. 饲粮添加木薯渣对羔羊生长性能、血清指标及瘤胃发酵指标的影响[J]. 动物营养学报, 2017, 29(10): 3666-3675. DOI:10.3969/j.issn.1006-267X.2017.10.029 |
| [8] |
丁冬梅. 浅谈泌乳奶牛不同阶段饲养管理技术[J]. 中国畜禽种业, 2015, 11(11): 99-100. DOI:10.3969/j.issn.1673-4556.2015.11.067 |
| [9] |
杨红建, 宋正河, 祝仕平.一种发酵微量气体产生量数据自动采集存储装置及方法: 中国, 200610011301.X[P].2007-12-19.
|
| [10] |
MENKE K H, STEINGASS H. Estimation of the energetic feed value obtained from chemical analysis and in vitro gas production using rumen fluid[J]. Animal Research and Development, 1988, 28: 7-55. |
| [11] |
ØRSKOV E R, MCDONALD I. The estimation of protein degradability in the rumen from incubation measurements weighted according to rate of passage[J]. The Journal of Agricultural Science, 1979, 92(2): 499-503. DOI:10.1017/S0021859600063048 |
| [12] |
VERDOUW H, VAN ECHTELD C J A, DEKKERS E M J. Ammonia determination based on indophenol formation with sodium salicylate[J]. Water Research, 1978, 12(6): 399-402. DOI:10.1016/0043-1354(78)90107-0 |
| [13] |
MAKKAR H P S, SHARMA O P, DAWRA R K, et al. Simple determination of microbial protein in rumen liquor[J]. Journal of Dairy Science, 1982, 65(11): 2170-2173. DOI:10.3168/jds.S0022-0302(82)82477-6 |
| [14] |
ERWIN E S, MARCO G J, EMERY E M. Volatile fatty acid analyses of blood and rumen fluid by gas chromatography[J]. Journal of Dairy Science, 1961, 44(9): 1768-1771. DOI:10.3168/jds.S0022-0302(61)89956-6 |
| [15] |
高红, 郝小燕, 张广宁, 等. 应用体外产气法和尼龙袋法评价几种粮食加工副产物的营养价值[J]. 中国饲料, 2017(7): 14-19. |
| [16] |
王芳, 徐元君, 牛俊丽, 等. 体外产气法评价反刍动物饲料营养价值的研究[J]. 中国畜牧兽医, 2016, 43(1): 76-83. |
| [17] |
王珊, 史良峰, 张振斌, 等. 麻叶荨麻和羊草比例对体外培养瘤胃微生物发酵参数的影响[J]. 家畜生态学报, 2018, 191(4): 39-43. |
| [18] |
NSAHLAI I V, SIAW D, OSUJI P O. The relationships between gas production and chemical composition of 23 browses of the genus Sesbania[J]. Journal of the Science of Food and Agriculture, 1994, 65(1): 13-20. DOI:10.1002/jsfa.2740650104 |
| [19] |
ASSOUMANI M B, VEDEAU F, JACQUOT L, et al. Refinement of an enzymatic method for estimating the theoretical degradability of proteins in feedstuffs for ruminants[J]. Animal Feed Science and Technology, 1992, 39(3/4): 357-368. |
| [20] |
卜登攀, 卢德勋, 崔慰贤, 等. 瘤胃能氮同步释放对瘤胃微生物蛋白质合成的影响[J]. 中国畜牧兽医, 2008, 35(12): 5-10. |
| [21] |
张成喜, 孙友德, 刘锡武, 等. 过瘤胃蛋氨酸对奶牛瘤胃微生物蛋白产量、产奶性能和氮排泄的影响[J]. 动物营养学报, 2017, 29(5): 1759-1766. |
| [22] |
马慧, 苗树君, 刘君. 反刍动物日粮正组合效应的调控及优化配制[J]. 中国牛业科学, 2005, 31(6): 47-50. DOI:10.3969/j.issn.1001-9111.2005.06.016 |
| [23] |
MATRAS J, BARTLE S J, PRESTON R L. Nitrogen utilization in growing lambs:effects of grain (starch) and protein sources with various rates of ruminal degradation[J]. Journal of Animal Science, 1991, 69(1): 339-347. DOI:10.2527/1991.691339x |
| [24] |
郭冬生, 彭小兰. 反刍动物尿素利用情况的探讨[J]. 湖南饲料, 2001(4): 24-25. |
| [25] |
王嘉厚.氨化稻草替代羊草对羔羊育肥效果及瘤胃发酵的影响[D].硕士学位论文.哈尔滨: 东北农业大学, 2009. http://www.wanfangdata.com.cn/details/detail.do?_type=degree&id=D072538
|
| [26] |
刘春龙, 姜文博, 付强, 等. 大豆黄酮和染料木素对奶牛瘤胃发酵的影响[J]. 中国畜牧杂志, 2011, 47(1): 60-63. |
| [27] |
BERGMAN E N. Glucose metabolism in ruminants as related to hypoglycemia and ketosis[J]. Cornell Veterinarian, 1973, 63(3): 341-382. |
| [28] |
冯仰廉. 反刍动物营养学[M]. 北京: 科学出版社, 2004: 335-336.
|
| [29] |
WHITELAW F G, EADIE J M, BRUCE L A, et al. Methane formation in faunated and ciliate-free cattle and its relationship with rumen volatile fatty acid proportions[J]. British Journal of Nutrition, 1984, 52(2): 261-275. DOI:10.1079/BJN19840094 |
| [30] |
熊本海, 卢德勋, 张子仪. 瘤胃乙酸与丙酸摩尔比例的改变对瘤胃发酵及血液指标的影响[J]. 畜牧兽医学报, 2002, 33(6): 537-543. DOI:10.3321/j.issn:0366-6964.2002.06.005 |
| [31] |
任莹, 赵胜军. 异位酸影响反刍动物瘤胃代谢的研究进展[J]. 饲料研究, 2008(2): 10-12. |
| [32] |
张慧玲, 李林, 陈勇, 等. 添加支链氨基酸或支链脂肪酸对玉米秸秆体外瘤胃发酵和细菌多样性的影响[J]. 新疆农业大学学报, 2012, 35(4): 265-269. DOI:10.3969/j.issn.1007-8614.2012.04.002 |