




2. 广州飞禧特生物科技有限公司, 广州 510640
2. Guangzhou Fishtech Biotechnology Co., Ltd., Guangzhou 510640, China
我国是水产养殖大国,养殖产量多年位居全球首位。饲料蛋白质是水产动物的主要能源物质。我国水产饲料工业使用的蛋白质资源(如鱼粉)较长时间处于短缺状态,依赖进口程度高,这制约着我国水产饲料工业和养殖业的可持续发展。但是,随着我国水产养殖业的高速发展,产业模式逐渐向高密度、集约化精养转变,养殖过程废水增多,易造成水体污染,破坏生态环境,不利于生态文明建设。“节能减排”一直是水产养殖业关注的方向。低蛋白质水平饲料(也称平衡氨基酸饲料)技术具有减少蛋白质原料的添加量、减少氮排放、改善动物生长性能的优势[1],是一种“节能减排”的有效手段。赵卿尧等[1]表明,低蛋白质水平饲料技术已经在畜禽养殖,特别是在猪生产中有了较深入的研究[2-3],在禽类[4]中也有了进一步的研究。在水产养殖中,低蛋白质水平饲料技术也有一定的开展,适量降低饲料中蛋白质含量不仅能提高草鱼(Ctenopharyngodon idellus)幼鱼生长性能,提高草鱼幼鱼的干物质和粗蛋白质表观消化率,还能减少草鱼幼鱼对水体的氮排泄[5]。低蛋白质水平饲料中添加色氨酸可提高凡纳滨对虾(Litopenaeus vannamei)饲料蛋白质、氨基酸、干物质和能量的表观消化率,提高蛋白酶活性,并改善全虾氨基酸的组成[6]。而陈乃松等[7]发现,在低蛋白质水平饲料中补充的晶体必需氨基酸对大口黑鲈(Largemouth bass)的生长、体组成和免疫指标产生的有益作用不及等量的以蛋白质为来源的必需氨基酸。因此,低蛋白质水平饲料技术在水产中的应用有待进一步研究。
核苷酸包括嘌呤核苷酸和嘧啶核苷酸,是动物体重要的组成成分,在机体能量转化、蛋白质合成、遗传信息编码、信号传导等几乎所有生物化学过程中都发挥重要作用。在机体处于快速生长、应激和病理状况下,机体快速分裂的免疫细胞、肠细胞等需要消耗大量的核苷酸,自身合成不足,需要外源补充,被称为“条件必需营养素”[8]。最初,核苷酸在水产动物中被作为一种诱食剂[9],而且混合核苷酸的诱食效果比单一核苷酸的好[10]。前人研究发现,饲料中添加一定量的核苷酸(或核苷酸类产品)具有促进水产动物生长[11-12]、提髙消化酶活性[13-14]、促进营养物质沉积[15]、提高免疫抗氧化能力[16-17]等作用。这表明核苷酸作为一种新型绿色添加剂具有保护水产动物健康和提高生产效益的作用。
饲料的营养价值由多种因素决定,包括饲料营养成分及动物对饲料中营养成分的消化、吸收和利用率等。表观消化率对评估饲料营养价值具有重要意义,是科学配制营养均衡饲料的前提。但是有关核苷酸对动物饲料营养物质表观消化率影响的报道尚不多见。饲料中添加0.2%核苷酸能提高断奶仔猪的粗脂肪、总磷、干物质和总能的表观消化率[18]。饲料中添加0.62%核苷酸能显著提高凡纳滨对虾对干物质和粗脂肪的表观消化率[19]。目前,水产动物低蛋白质水平饲料中添加核苷酸对营养物质表观消化率的研究尚未见报道。因此,本试验旨在探究核苷酸对凡纳滨对虾低蛋白质水平饲料的干物质、粗蛋白质、粗脂肪、总磷、氨基酸和脂肪酸的表观消化率的影响,以期为核苷酸在对虾低蛋白质水平饲料中的应用提供参考数据。
1 材料与方法 1.1 试验材料5′-胞苷酸、5′-腺苷酸、5′-肌苷酸二钠、5′-尿苷酸二钠和5′-鸟苷酸二钠由南京某生物技术有限责任公司提供,纯度均>99%。试验所用核苷酸为上述5种核苷酸按质量比为1 : 1 : 1 : 1 : 1混合而成的核苷酸混合物。
1.2 试验设计与饲料组成本试验设2个对照组,低蛋白质对照组饲料粗蛋白质水平为37.92%,粗脂肪水平为6.74%,记为L0组;高蛋白质对照组饲料粗蛋白质水平为46.45%,粗脂肪水平为6.57%,记为H0组;此外,在L0组基础上添加400 mg/kg核苷酸混合物,记为LN组。3种试验饲料按照等脂方式配制。
饲料原料粉碎过80目筛,微量成分采用逐级扩大法混匀后,再加入油和水,用双螺杆压机(SLX-80型)制得直径2.0 mm颗粒料,经55 ℃烘干7~8 h,自然冷却,过筛装袋,室温保存备用。各组饲料组成及营养水平、饲料氨基酸的组成、饲料脂肪酸的组成如表 1、表 2、表 3所示。
|
|
表 1 饲料组成及营养水平(风干基础) Table 1 Composition and nutrient levels of diets (air-dry basis) |
|
|
表 2 饲料的氨基酸组成(风干基础) Table 2 Amino acid composition of diets (air-dry basis) |
|
|
表 3 饲料的脂肪酸组成(风干基础) Table 3 Fatty acid composition of diets (air-dry basis) |
凡纳滨对虾虾苗购自广东省中山市合一虾苗场。先以商品饲料在室内循环水养殖系统进行7 d暂养,每天分别在08:00、14:00和20:00分3次投喂。室内循环水养殖系统由24个玻璃纤维桶构成,其容积为350 L(直径80 cm,高70 cm,实际水量300 L),进水速率为2.6~3.2 L/min。正式试验在广东省农业科学院动物科学研究所室内循环水养殖系统内进行。挑选健康且体质量为(2.00±0.01) g凡纳滨对虾480尾,随机分成3组,每组4个重复,每个重复40尾。各组分别投喂3种试验饲料。采用表观饱食投喂法,每天在08:00、14:00、20:00进行投喂。每次投喂30 min后采取虹吸法清理残饵和粪便。正式投喂后的第10天开始,每次投喂1 h后通过虹吸法收集完整的粪便,连续收集30 d。收集的各重复虾粪便置于60 ℃烘箱中烘干后,-20 ℃冰箱中保存备用。将消毒过的海水和曝气的自来水混合,经砂滤后供养殖过程使用。每3~4 d吸污1次,吸污后换1/3水。每天记录对虾的死亡情况。每周测定水质情况。自然光照。养殖过程中水盐度4.5~5.5,水温24.4~29.5 ℃,pH 7.7~8.0,溶氧含量>5.0 mg/L,氨氮含量≤0.1 mg/L,亚硝酸盐含量≤0.01 mg/L。
1.4 检测指标和计算方法饲料原料及饲料和粪便中水分、粗蛋白质、粗脂肪、粗灰分和总磷含量分别按照GB/T 5009.3—2016、GB/T 6432—2018、GB/T 6433—2006、GB/T 6438—2007、GB/T 6437—2018的方法进行测定。饲料和粪便的氨基酸组成和含量按照GB/T 18246—2000的方法,使用安捷伦1260型液相色谱仪通过衍生法测定。饲料和粪便通过甲酯化前处理,使用安捷伦6890型气相色谱仪按照GB/T 17377—2008的方法测定脂肪酸组成和含量。饲料和粪便样品中钇含量委托广州分析测试中心采用等离子发光色谱仪进行测定。
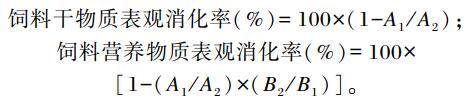
|
式中:A1为饲料中三氧化二钇(Y2O3)含量;A2为粪便中Y2O3含量;B1为饲料中某种营养成分含量;B2为粪便中某种营养成分含量。
1.5 数据统计试验数据用SPSS 17.0软件进行单因素方差分析(one-way ANOVA),结果采用平均值±标准差显示,若组间差异显著,再用Duncan氏法进行多重比较,以P < 0.05为差异显著标准。
2 结果 2.1 各组干物质、粗蛋白质、粗脂肪、粗灰分和总磷表观消化率由表 4可知,H0组的干物质和粗蛋白质表观消化率最低,L0组其次,LN组最高,各组之间无显著差异(P>0.05)。LN组的粗脂肪和总磷表观消化率显著高于H0组(P<0.05),与L0组无显著差异(P>0.05)。LN组的粗灰分表观消化率最高,与L0和H0组无显著差异(P>0.05)。
|
|
表 4 核苷酸对凡纳滨对虾饲料中干物质、粗蛋白质、粗脂肪、粗灰分和总磷表观消化率的影响 Table 4 Effects of nucleotide on apparent digestibilities of dry matter, crude protein, ether extract, ash and total phosphorus in diets of Litopenaeus vannamei (n=4) |
由表 5可知,各组之间的各氨基酸、非必需氨基酸和必需氨基酸表观消化率均无显著差异(P>0.05)。其中,LN组的天冬氨酸表观消化率最低,H0组最高。LN组的谷氨酸、丝氨酸、甘氨酸、丙氨酸和非必需氨基酸表观消化率高于H0组,但均低于L0组。LN组的酪氨酸表观消化率高于L0和H0组。LN组的苏氨酸、组氨酸、蛋氨酸表观消化率低于L0和H0组。LN组的精氨酸、缬氨酸、苯丙氨酸、亮氨酸和必需氨基酸表观消化率高于H0组,但均低于L0组。LN组的异亮氨酸和赖氨酸表观消化率高于H0和L0组。
|
|
表 5 核苷酸对凡纳滨对虾饲料中氨基酸表观消化率的影响 Table 5 Effects of nucleotide on apparent digestibilities of amino acids in diets of Litopenaeus vannamei (n=4) |
由表 6可知,各组之间的单不饱和脂肪酸、多不饱和脂肪酸、总脂肪酸表观消化率无显著差异(P>0.05)。L0组的C12 : 0、C15 : 0、C17 : 0、isoC19 : 0、C20 : 0和C22 : 0表观消化率均高于H0组(P>0.05),L0组的C14 : 0表观消化率显著高于H0组(P<0.05)。LN组的饱和脂肪酸、C17 : 0、C20 : 0表观消化率均显著高于L0和H0组(P<0.05),但L0组和H0组之间无显著差异(P>0.05)。LN组的C16 : 1表观消化率显著高于H0组(P<0.05),LN组的C22 : 1、C18 : 3和DHA表观消化率显著高于L0和H0组(P<0.05),但L0组和H0组之间无显著差异(P>0.05)。
|
|
表 6 核苷酸对凡纳滨对虾饲料中脂肪酸表观消化率的影响 Table 6 Effects of nucleotide on apparent digestibilities of fatty acids in diets of Litopenaeus vannamei (n=4) |
干物质表观消化率是饲料中营养物质(包括粗蛋白质、粗脂肪、粗灰分、总磷等)总体消化水平的体现。在本试验中,与高、低蛋白质水平饲料相比,虽然在低蛋白质水平饲料中添加核苷酸能提高凡纳滨对虾对饲料中干物质表观消化率,但效果不明显。这与前人研究结果有所不同。周乐等[18]研究发现,饲料中添加0.05%和0.10%核苷酸能提高断奶仔猪干物质表观消化率,而当核苷酸添加量为0.20%时,其干物质表观消化率显著提高。Li等[20]也报道,在7~42日龄仔猪饲粮中添加核苷酸能提高仔猪的干物质、粗蛋白质和总能的表观消化率,提高仔猪的生长性能。仔猪饲粮中添加0.5%酵母核苷酸产品能显著提高其干物质和粗蛋白质表观消化率[21]。饲料中添加0.62%核苷酸混合物能显著提高凡纳滨对虾干物质表观消化率[22]。核苷酸能提高动物对干物质表观消化率原因,Li等[20]报道认为是核苷酸可通过促进提高水产动物的摄食速率,减少饲料中营养物质的溶失,更多用于体内的代谢过程,促进营养物质的沉积,从而提高动物生长。但也有不同研究结果,饲料中添加核苷酸对草鱼(Ctenopharyngodon idellus)[16]、尼罗罗非鱼(Oreochromis niloticus×O.areus)[23]全鱼的干物质(或水分)含量无显著效果。这可能与核苷酸对干物质表观消化率与饲料原料组成、营养水平及核苷酸类型和用量有关,此外,动物种类、生长阶段以及养殖环境等也会对干物质表观消化率有所影响,具体原因有待进一步探讨。
3.2 核苷酸对凡纳滨对虾饲料中蛋白质及氨基酸表观消化率的影响蛋白质表观消化率是评估动物对饲料蛋白质消化利用程度的重要指标。饲料蛋白质水平、氨基酸组成和含量、机体对蛋白质的消化吸收能力都与蛋白质的消化率密切相关。氨基酸是蛋白质的基本组成,对蛋白质的消化实质是对氨基酸的利用。本试验结果显示,饲料中添加核苷酸对凡纳滨对虾饲料粗蛋白质、6种非必需氨基酸、9种必需氨基酸、总非必需氨基酸和总必需氨基酸的表观消化率均无显著影响,这与饲料中添加核苷酸对断奶仔猪的粗蛋白质表观消化率无显著影响的结果类似[18]。而陈冰等[19]研究发现,在植物蛋白质源替代鱼粉饲料中添加0.62%核苷酸混合物对凡纳滨对虾的粗蛋白质表观消化率无显著影响,但能显著改善凡纳滨对虾对饲料的苯丙氨酸、组氨酸、异亮氨酸、亮氨酸、苏氨酸、缬氨酸、蛋氨酸、总必需氨基酸、天门冬氨酸、脯氨酸、谷氨酸、甘氨酸、丙氨酸、半胱氨酸和总非必需氨基酸的表观消化率。核苷酸对蛋白质表观消化率研究结果迥异,与核苷酸产品来源、核苷酸单体混合物种的各成分的含量、养殖对象、规格和所处的养殖环境、养殖周期和核苷酸添加方式都有一定的关系。此外,核苷酸对不同动物影响作用途径,也会对蛋白质表观消化吸收率产生影响。饲料中添加混合核苷酸能提高大黄鱼血液中血红蛋白和总蛋白含量,但是差异不显著[17]。据报道,饲料中添加0.03%的核苷酸对凡纳滨对虾血清中的总蛋白含量影响不显著[24]。王锐等[25]研究发现,在异育银鲫(Carassius auratus gibelio)幼鱼饲料中添加核苷酸能提高其肠道和肝胰腺消化酶活性。但是也有一些研究存在不同的结果,饲料中添加适量的核苷酸显著提高斑点叉尾(Ictalurus punctatus)[26]鱼体粗蛋白质含量。饲料中添加核苷酸能显著提高半滑舌鳎(Cynoglossus semilaevis)稚鱼的胰脏和肠道蛋白酶活性[13]。饲料中添加0.06%核苷酸显著提高了草鱼的肌蛋白和中肠总蛋白含量,提高了肝蛋白和前肠总蛋白含量[27]。饲料中添加1.5 g/kg的核苷酸能显著提高黄颡鱼的蛋白质沉积率[28]。昝林森等[29]推测核苷酸可能是通过作为蛋白激酶促进代谢酶和激素的生成,进而促进蛋白质的合成。
3.3 核苷酸对凡纳滨对虾饲料中粗脂肪和脂肪酸表观消化率的影响脂肪是动物组织中主要的组成物质和储能物质,为机体提供能量和脂肪酸。高等动物中已有核苷酸对脂肪代谢调节作用的研究。在奶粉配方中添加核苷酸能够提高早产儿肠上皮细胞、足月婴儿肠上皮细胞和血浆中长链多不饱和脂肪酸的含量。关于饲料中添加核苷酸对动物饲料中粗脂肪表观消化率的影响,前人已有少量研究报道。周乐等[18]研究发现,饲料中添加核苷酸能显著提高断奶仔猪的粗脂肪表观消化率。陈冰等[19]也发现核苷酸能显著提高凡纳滨对虾对饲料中粗脂肪表观消化率。在真鲷(Pagrus major)幼鱼饲料中添加0.2%腺嘌呤核糖核苷酸能提高粗脂肪表观消化率[30]。本试验结果与上述结果相似,但本试验结果还发现饲料中添加核苷酸能显著提高饱和脂肪酸、C17 : 0、C20 : 0、C22 : 1、C18 : 3、DHA表观消化率,而C14 : 0、C16 : 1表观消化率显著高于H0组,与L0组无显著差异。近年来有关核苷酸对水产动物脂代谢的作用受到关注。李解[31]发现添加0.1%复合核苷酸在促进斑马鱼(Danio rerio)生长的同时可以降低肝脏脂肪含量并提高肌肉脂肪含量。饲料中添加混合核苷酸能显著提高大黄鱼血液中甘油三酯和胆固醇含量[17]。已有研究发现,饲料中添加核苷酸能显著提高鲟鱼(Beluga sturgeon)的鱼体饱和脂肪酸和不饱和脂肪酸含量,而各组间的单不饱和脂肪酸、多不饱和脂肪酸、n-3多不饱和脂肪酸、n-6多不饱和脂肪酸、n3/n6和二十碳五烯酸+二十二碳六烯酸含量差异不显著[32]。饲料中添加核苷酸能显著提高凡纳滨对虾的虾体C14 : 1n-5、C20 : 3n-3和C22 : 6n-6含量,但对饱和脂肪酸、单不饱和脂肪酸以及多不饱和脂肪酸含量没有显著影响,推测这可能是脂肪酶对不同的脂肪酸的消化吸收能力不一致导致的[33]。不同的研究对象及脂肪酸的长度、饱和度、组成和含量等都能影响机体对脂肪酸的消化吸收能力,进而影响机体对粗脂肪的消化能力。核苷酸对脂肪代谢影响的作用机理还比较缺乏,其具体机制并不明确。
4 结论与高、低蛋白质水平饲料相比,在低蛋白质水平饲料中添加核苷酸具有改善凡纳滨对虾对饲料营养物质表观消化率的效果,特别是对粗脂肪、总磷和脂肪酸。
| [1] |
赵卿尧, 顾宪红. 低蛋白日粮应用前景与研究进展[J]. 家畜生态学报, 2020, 41(3): 9-14. |
| [2] |
于树龙.低蛋白日粮平衡氨基酸对生长猪生长性能及氮排放的影响研究[D].硕士学位论文.武汉: 华中农业大学, 2015.
|
| [3] |
贾晨晨.低蛋白日粮添加不同蛋白酶对育肥猪生长性能和养分表观消化率的影响[D].硕士学位论文.郑州: 河南农业大学, 2015.
|
| [4] |
WANG Q D, ZHANG K Y, ZHANG Y, et al. Effects of dietary protein levels and protease supplementation on growth performance, carcass traits, meat quality, and standardized ileal digestibility of amino acid in pekin ducks fed a complex diet[J]. Poultry Science, 2020, 99(7): 3557-3566. DOI:10.1016/j.psj.2020.03.047 |
| [5] |
刘梦梅.低蛋白饲料中添加赖氨酸和蛋氨酸对草鱼幼鱼生长及氮排泄的影响[D].硕士学位论文.武汉: 华中农业大学, 2016.
|
| [6] |
孙育平, 裘金木, 王国霞, 等. 低蛋白质饲料中添加色氨酸对凡纳滨对虾饲料表观消化率、消化酶活和全虾氨基酸组成的影响[J]. 水生生物学报, 2016, 40(4): 720-727. |
| [7] |
陈乃松, 梁勤朗, 肖温温, 等. 在低蛋白质饲料中补充必需氨基酸对大口黑鲈生长、体组成和免疫指标的影响[J]. 水生生物学报, 2014, 38(2): 262-271. |
| [8] |
MATEO C D, STEIN H H. 核苷酸对幼龄动物肠道发育和免疫功能的影响[J]. 饲料工业, 2014, 35(16): 58-64. |
| [9] |
赵红月.异育银鲫促摄食物质研究[D].博士学位论文.武汉: 中国科学院研究生院(水生生物研究所), 2007.
|
| [10] |
LI P, LAWRENCE A L, CASTILLE F L, et al. Preliminary evaluation of a purified nucleotide mixture as a dietary supplement for Pacific white shrimp Litopenaeus vannamei (Boone)[J]. Aquaculture Research, 2007, 38(8): 887-890. DOI:10.1111/j.1365-2109.2007.01761.x |
| [11] |
苗新, 曹娟娟, 徐玮, 等. 核苷酸对大黄鱼生长性能、肠道形态和抗氧化能力的影响[J]. 水产学报, 2014, 38(8): 1140-1148. |
| [12] |
许丹丹, 曹俊明, 黄燕华, 等. 饲料中添加核苷酸对凡纳滨对虾幼虾生长、肠道形态及抗氧化酶活力的影响[J]. 中国水产科学, 2011, 18(5): 1115-1124. |
| [13] |
刘经纬.饲料中谷氨酰胺和核苷酸对半滑舌鳎稚鱼生长、消化酶活力、抗氧化酶活力及基因表达的影响[D].硕士学位论文.青岛: 中国海洋大学, 2015.
|
| [14] |
刘扬, 冯琳, 赵波, 等. 豆粕替代部分鱼粉饲料中添加核苷酸和有机硒对幼建鲤生长和消化吸收功能的影响[J]. 四川农业大学学报, 2012, 30(2): 238-242. |
| [15] |
曹俊明, 许丹丹, 黄燕华, 等. 饲料中添加核苷酸对凡纳滨对虾幼虾生长、组织生化组成及非特异性免疫功能的影响[J]. 水产学报, 2011, 35(4): 594-603. |
| [16] |
王红权, 李瑞, 金柏涛, 等. 饲料中添加核苷酸对草鱼生长性能、体成分及血清生化指标的影响[J]. 动物营养学报, 2019, 31(6): 2745-2751. |
| [17] |
吴文俊, 周飘苹, 黎明, 等. 饲料中添加不同核苷酸对大黄鱼生长、血液指标及血清酶活性的影响[J]. 宁波大学学报(理工版), 2014, 27(2): 7-12. |
| [18] |
周乐, 何进田, 徐稳, 等. 核苷酸对断奶仔猪生长性能、养分表观消化率和血液指标的影响[J]. 家畜生态学报, 2017, 38(3): 46-51. |
| [19] |
陈冰, 齐飞, 黄燕华, 等. 豆粕和花生粕部分替代鱼粉添加四种添加剂对凡纳滨对虾饲料表观消化率的影响[J]. 饲料工业, 2014, 35(20): 19-24. |
| [20] |
LI H L, ZHAO P Y, LEI Y, et al. Response to an Escherichia coli K88 oral challenge and productivity of weanling pigs receiving a dietary nucleotides supplement[J]. Journal of Animal Science and Biotechnology, 2015, 6: 49. DOI:10.1186/s40104-015-0049-5 |
| [21] |
KIM S W.酵母培养物对保育猪的生长性能、肠道健康和免疫应答的影响[J].郭添福译.江西饲料, 2010(1):27-35.
|
| [22] |
LI P, GATLIN Ⅲ D M. Nucleotide nutrition in fish:current knowledge and future applications[J]. Aquaculture, 2005, 251(2/3/4): 141-152. |
| [23] |
韩春艳, 郑清梅, 邓茜壬. 外源核苷酸对罗非鱼生长发育及抗氧化能力的影响[J]. 嘉应学院学报, 2018, 36(5): 39-44. |
| [24] |
胡俊茹, 曹俊明, 黄燕华, 等. 几种诱食剂对凡纳滨对虾生长、血清生化指标和肝胰腺消化酶的影响[J]. 淡水渔业, 2010, 40(2): 30-35. |
| [25] |
王锐, 王春维, 李星燕, 等. 外源核苷酸对异育银鲫幼鱼消化酶活性的影响[J]. 中国水产, 2008(3): 70-72. |
| [26] |
沙航, 王锐, 祝爱侠, 等. 核苷酸对斑点叉尾鮰幼鱼生长、体成分和胃肠道形态的影响[J]. 粮食与饲料工业, 2015(2): 44-48. |
| [27] |
唐德约.饲料中添加核苷酸对草鱼生长性能及肠道的影响[D].硕士学位论文.长沙: 湖南农业大学, 2014.
|
| [28] |
赵红霞, 曹俊明, 黄燕华, 等.饲料中添加核苷酸对黄颡鱼生长性能、生理指标和抗氧化功能的影响[C]//第九届世界华人鱼虾营养学术研讨会论文集.厦门: 中国水产学会, 2013.
|
| [29] |
昝林森, 邱怀. 关于环核苷酸与家畜生长之调控[J]. 甘肃畜牧兽医, 1995(5): 27-28. |
| [30] |
HOSSAIN S, KOSHIO S, ISHIKAWA M, et al. Dietary effects of adenosine monophosphate to enhance growth, digestibility, innate immune responses and stress resistance of juvenile red sea bream, Pagrus major[J]. Fish & Shellfish Immunology, 2016, 56: 523-533. |
| [31] |
李解.外源核苷酸对斑马鱼脂代谢和免疫的调控和相关机理研究[D].硕士学位论文.北京: 中国农业科学院, 2019.
|
| [32] |
ABTAHI B, YOUSEFI M, KENARI A A. Influence of dietary nucleotides supplementation on growth, body composition and fatty acid profile of Beluga sturgeon juveniles (Huso huso)[J]. Aquaculture Research, 2011, 44(2): 254-260. |
| [33] |
OUJIFARD A, KENARI A A.Growth, lipid metabolism and intestinal absorbance of white shrimp (Litopenaeus Vannamei Boone 1931) influenced by dietary nucleotide[C]//2nd International Conference on Environment, Agriculture and Food Sciences (ICEAFS'2013).Kuala Lumpur: Malaysia, 2013.
|