




能量类饲料在毛皮动物饲粮中占较大比例[1],确定水貂能量需要量,使其获得良好的生产性能,减少饲粮浪费,是高效养殖的关键。目前,国内主要参考NRC(1982)[2]和NJF(2012)[3]营养标准进行毛皮动物各生长阶段饲粮的配制,但不同国家动物品种、饲养管理、饲粮结构及组成存在差异,上述营养标准并不完全适用于我国毛皮动物养殖需求;且至今毛皮动物能量评定多使用代谢能(ME)体系,毛皮动物净能(NE)体系的研究相对较少。NE是ME与产热量(HP)的差值,较ME更加准确地反映动物能量需要量[4-5]。温度是影响动物HP的重要因素[6-7],水貂冬毛生长期间正处于东北地区气候严寒、温度低的时期,而此时确是水貂毛皮生长的重要阶段,毛皮品质是影响毛皮动物养殖经济效益的主要因素[8]。因此,本试验在本课题组已确定冬毛生长期雄性水貂适宜ME和粗蛋白质水平[9]的基础上,使用开放回流式呼吸测热装置,研究冬毛生长期雄性水貂NE需要量,为我国水貂生产提供科学准确的参考数据。
1 材料与方法 1.1 试验动物与设计选取(145±5)日龄、体重[(1 823.75±64.68) g]相近的冬毛生长期健康雄性短毛黑水貂30只,饲喂前期评估出的最佳ME与粗蛋白质水平饲粮,预饲7 d,使水貂适应试验饲粮后进行呼吸测热试验。呼吸测热试验共设3期,每期7 d,每期3个重复,每个重复2只水貂。每期呼吸测热试验开始前,将试验动物空腹称重并从中选取6只水貂,每2只放进呼吸测热装置的1个代谢室内,代谢室内连续测定气体交换7 d,其中适应期1 d,正常代谢3 d,绝食代谢3 d,正式试验时间为2018年11月7日至2018年11月28日。
1.2 试验饲粮依据NRC(1982)和NJF(2012)营养标准及国内相关研究,以膨化玉米粉、膨化大豆、豆粕、肉骨粉、玉米蛋白粉、鸡肉粉、鱼粉、豆油为主要原料,添加水貂生长发育所需的维生素和矿物质(由河北某农业有限公司提供)配制冬毛生长期雄性水貂饲粮。试验饲粮组成及营养水平见表 1。
|
|
表 1 试验饲粮组成及营养水平(风干基础) Table 1 Composition and nutrient levels of the experimental diet (air-dry basis) |
动物饲养试验在农业农村部长白山野生生物资源重点野外科学观测站开展,试验动物由专人饲养。呼吸测热装置呼吸小室内设温度为15 ℃,正常采食期间每天08:00—10:00进行动物饲喂、称重和采样等工作,绝食代谢期间保证自由饮水。
1.4 呼吸测热装置该试验所用的6室并联毛皮动物开放回流式呼吸测热装置由吉林省农业科学院杨华明研究员团队研制。该套装置共有6个代谢室,代谢室体积为0.42 m3(100 cm×70 cm×60 cm),代谢室内设有饮水装置,粪、尿收集装置以及气体循环、制冷、加热等设备。数据采集控制仪按照试验流程驱动气体分析仪传感器依次对户外空气和代谢室(A~F室)按先后顺序循环采集,循环切换时间自行设定为3 min,软件自动计算水貂消耗氧气(O2)消耗量、二氧化碳(CO2)产生量、呼吸熵(RQ),记录代谢室内的温度和湿度。
1.5 样品及数据收集呼吸测热试验期内,第1天水貂置于呼吸室内适应小室环境。第2天饲喂前空腹称重并记录数据;第2~4天正常饲喂,记录采食量;每天08:00—10:00收集粪样、尿样,并提前在尿盒内加入5 mL 10%硫酸,连续收集尿液和粪便3 d,置于-20 ℃保存待测。第5天早上再次空腹称重,第5~7天水貂进行绝食代谢,连续收集尿液,第8天早上试验动物再次空腹称重。整个呼吸测热周期内连续测定水貂O2消耗量、CO2产生量和RQ。
1.6 测定指标及方法参考张丽英[10]主编的《饲料分析及饲料质量检测技术》中的方法对试验饲粮、粪便和尿液进行化学成分分析。饲粮、粪便样品的干物质含量通过在105 ℃干燥至恒重的方法测定;用全自动凯氏定氮仪(FOSS Kjeltec-8400,Hillerød,丹麦)测量氮含量;以苯甲酸为标准品,用氧弹测热仪(IKAC-2000, Calorimeter, IKA company, 德国)测定饲粮和粪便的总能。采用滤纸法测定尿能,取10张直径为9 cm的定量滤纸,每2张共同折叠称重,计算5组滤纸的总能平均值。将7 mL尿液滴在2张滤纸上置于特制坩埚中,在65 ℃干燥8 h,用氧弹测热仪测定尿液总能。
水貂HP根据Brouwer[11]的公式计算:
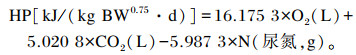
|
代谢能摄入量(MEI)与沉积能(RE)建立的线性回归方程[12-13]:
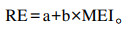
|
式中:a、b为常数。
MEI与HP建立的线性回归方程[14]:
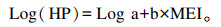
|
式中:a为绝食产热量(FHP);b为常数。
1.7 数据分析及处理使用Excel 2016处理原始数据,采用SAS 9.2软件进行平均值和标准误统计分析,采用PROC REG模型对MEI与RE,MEI与HP反对数进行线性回归分析,试验结果以“平均值±标准误”表示,P<0.05为差异显著,P<0.01为差异极显著,P>0.05为差异不显著。
2 结果 2.1 冬毛生长期雄性水貂能量平衡利用呼吸测热装置检测分析,水貂正常采食时MEI为755.69 kJ/(kg BW0.75·d),HP为526.31 kJ/(kg BW0.75·d),生长净能(NEg)为229.00 kJ/(kg BW0.75·d);FHP为489.75 kJ/(kg BW0.75·d),NE需要量为718.75 kJ/(kg BW0.75·d),具体结果见表 2。
|
|
表 2 冬毛生长期雄性水貂能量平衡参数 Table 2 Energy balance parameters of male minks during winter fur-growing period |
通过开放式呼吸测热装置测得,冬毛生长期水貂正常采食状态时O2消耗量为26.19 L/(kg BW0.75·d),CO2产生量为21.26 L/(kg BW0.75·d),RQ为0.82。绝食状态时O2消耗量为24.63 L/(kg BW0.75·d),CO2产生量为19.44 L/(kg BW0.75·d),RQ为0.79。具体结果见表 3。
|
|
表 3 冬毛生长期雄性水貂呼吸代谢 Table 3 Respiratory metabolism of male minks during winter fur-growing period |
|
|
表 4 冬毛生长期雄性水貂维持净能与维持代谢能需要量(体重1.92~2.40 kg) Table 4 NEm and MEm requirements of male minks during winter fur-growing period (BW 1.92 to 2.40 kg) |
图 1、图 2显示,MEI与RE建立的线性回归方程为RE=-496.20(±11.413 7)+0.96(±0.014 9)MEI(R2=0.999 0, P < 0.000 1),经方程计算,当MEI为0时,得维持净能(NEm)为493.20 kJ/(kg BW0.75·d),当RE为0时,得维持代谢能(MEm)为519.80 kJ/(kg BW0.75·d),Km为95.46%。MEI与HP反对数建立的线性回归方程为Log(HP)=2.676 0(±0.006 7)+6.78×10-5(±0.000 008 7)MEI(R2=0.937 8, P=0.001 5),由方程计算,当MEI=0时,得NEm为474.24 kJ/(kg BW0.75·d),当MEI=HP时,得MEm为514.00 kJ/(kg BW0.75·d),Km为92.27%。

|
图 1 冬毛生长期雄性水貂MEI与RE的回归关系 Fig. 1 Relationship between MEI and RE of male minks during winter fur-growing period |

|
图 2 冬毛生长期雄性水貂MEI与HP反对数的回归关系 Fig. 2 Relationship between MEI and logarithm of HP of male minks during winter fur-growing period |
能量平衡是评价饲粮的重要指标,根据NJF(2012)营养标准,11月水貂MEI为602.56~796.88 kJ/(kg·d);杨颖[15]报道,饲粮ME水平为15.0~16.5 MJ/kg时,冬毛生长期雄性水貂MEI为899.52~1 165.74 kJ/(kg BW0.75·d),本试验结果与上述研究结果基本一致。温度是影响机体热增耗的重要因素,有文献报道,相比于20 ℃,当环境温度为28 ℃时,热增耗升高11.0~12.5 kJ/(kg BW0.75·d);在一定范围内随着畜舍中环境温度的降低,仔猪HP呈线性上升[16];外界温度低于18 ℃时,每降低1 ℃生长猪维持产热量则线性增加15.48~18.83 kJ/(kg BW0.75·d)[17]。当外界温度低于临界温度,水貂需要补充能量来维持机体代谢时,每降低1 ℃,HP会线性增加9.6~15.5 kJ/(kg·d)[1, 18-19]。本试验结果显示,冬毛生长期雄性水貂HP为526.31 kJ/(kg BW0.75·d),与Wamberg[20]研究结果501 kJ/(kg·d)基本接近。李昌锐[21]报道冬毛生长期水貂能量需要量较高,以储备脂肪维持体温,该时期RE低于育成生长期。杨嘉实等[22]报道冬毛生长期水貂FHP为559.12 kJ/(kg·d),高于育成生长期FHP[508.06 kJ/(kg·d)],本试验结果显示,水貂FHP为489.75 kJ/(kg BW0.75·d),高于育成生长期FPH[414.87 kJ/(kg BW0.75·d)],与上述结果趋势一致,但可能是由于饲粮配方、试验温度不同导致本试验FHP略低于上述研究。目前国内外缺乏水貂NE需要量研究及相关数据,本试验结果显示冬毛生长期水貂NE需要量为718.75 kJ/(kg BW0.75·d),为水貂养殖生产及营养标准的制定提供数据参考。
3.2 冬毛生长期雄性水貂呼吸代谢通过呼吸测热装置检测动物气体交换过程中O2消耗量和CO2产生量,可以间接计算水貂的HP[23-24]。研究报道,水貂较普通畜禽对脂肪需要量较高[25],冬毛生长期水貂MEI较低时,机体会降低碳水化合物、蛋白质供能所占比例,脂肪供能大大增加[26],以维持正常机体代谢和体温。本试验结果显示,正常采食期间水貂O2消耗量为26.19 kJ/(kg BW0.75·d)、CO2产生量为21.26 kJ/(kg BW0.75·d),且均高于绝食代谢。研究报道,自由采食状态下动物RQ为0.7~1.0,绝食状态下RQ为0.70以下[27]。本试验中,正常采食转为绝食代谢时水貂RQ由0.82降至0.79,比上述RQ略高,可能是由于物种差异,水貂比家禽储存的皮下脂肪多,绝食代谢时水貂主要氧化分解机体脂肪供能[28],饥饿耐受力较强。
3.3 冬毛生长期雄性水貂能量需要量钟伟等[1]报道,夏季天气炎热,水貂热量消耗较高;9~11月份则需要积累脂肪以度过寒冬,因此7~12月成年雄性水貂的MEm呈升高趋势。杨嘉实等[22]报道冬毛生长期水貂MEm为579.62 kJ/(kg BW0.75·d)。Harper等[29]表示当水貂体重为2.00 kg时,水貂MEm为519.56 kJ/(kg BW0.75·d)。本试验水貂体重为1.92~2.40 kg,由线性回归方程所得MEm为514.00~519.80 kJ/(kg BW0.75·d),NEm为474.24~493.20 kJ/(kg BW0.75·d);本试验结果与上述研究结果基本相近。杨嘉实[24]研究报道,冬毛生长期水貂Km为96.46%,本试验中由方程所得Km为92.27%~95.46%,与上述研究结果基本一致,该时期毛皮动物Km较高,可能是因为冬毛生长期较低的外界温度使得水貂采食活动减弱,采食量降低,营养物质代谢减弱,采食或绝食状态下机体产热主要用于维持基础代谢和较弱的采食活动,因此摄入的能量得到了充分的利用。
4 结论综上所述,体重为1.92~2.40 kg冬毛生长期雄性短毛黑水貂能量需要量为:MEm 514.00~519.80 kJ/(kg BW0.75·d),NEm 474.24~493.20 kJ/(kg BW0.75·d),NEg 229.00 kJ/(kg BW0.75·d),NE 718.75 kJ/(kg BW0.75·d);Km 92.27%~95.46%。
| [1] |
钟伟, 穆琳琳, 张婷, 等. 毛皮动物能量代谢的研究进展[J]. 动物营养学报, 2018, 30(8): 2879-2886. |
| [2] |
NRC.Nutrient requirements of mink and foxes[S]. 2nd ed.Washington, D.C.: National Academies Press, 1982.
|
| [3] |
NJF.Energy and main nutrients in feed for mink and foxes[S]. 2nd ed.Nykarleby (Finland): Fur Animals Nutrition and Feeding Committee, 2012: 12-15.
|
| [4] |
焦光月, 马雪豪, 金亚倩, 等. 肉用羊能量与蛋白质需要量测定方法及研究进展[J]. 饲料博览, 2015(6): 20-23. |
| [5] |
王旭莉.蛋鸡玉米和豆粕净能值的测定及其净能体系的应用[D].硕士学位论文.杨凌: 西北农林科技大学, 2010: 3-7.
|
| [6] |
王利华, 周丹, 王光. 环境温度对哈白生长猪代谢产热量的影响[J]. 家畜生态, 2002, 23(2): 47-48. |
| [7] |
常玉, 冯京海, 张敏红. 环境温度、湿度等因素对家禽体温调节的影响及评估模型[J]. 动物营养学报, 2015, 27(5): 1341-1347. |
| [8] |
穆琳琳, 钟伟, 陈双双, 等. 饲粮维生素B2添加水平对冬毛期水貂生长性能、毛皮品质、营养物质消化率及氮代谢的影响[J]. 动物营养学报, 2019, 31(1): 251-257. |
| [9] |
韩菲菲, 钟伟, 张新宇, 等. 饲粮代谢能和粗蛋白质水平对冬毛期雄性水貂生产性能、氮代谢及能量利用率的影响[J]. 动物营养学报, 2019, 31(11): 5157-5167. |
| [10] |
张丽英. 饲料分析及饲料质量检测技术[M]. 3版. 北京: 中国农业大学出版社, 2007: 126-127.
|
| [11] |
BROUWER E.Report of sub-committee on constants and factors[C]//Proceedings of the 3rd Symposium on Energy Metabolism.London (UK): Academic Press, 1965: 441-443.
|
| [12] |
FARREL D J.General principles and assumptions of calorimetry[M]//MORRIS T R, FREEMAN B M.Energy requirements of poultry.Edinburgh, UK: British Poultry Science, 1974: 1-23
|
| [13] |
SAKOMURA N K, LONGO F A, OVIEDO-RONDON E O, et al. Modeling energy utilization and growth parameter description for broiler chickens[J]. Poultry Science, 2005, 84(9): 1363-1369. |
| [14] |
LOFGREEN G P, GARRETT W N. A system for expressing net energy requirements and feed values for growing and finishing beef cattle[J]. Journal of Animal Science, 1968, 27(3): 793-806. DOI:10.2527/jas1968.273793x |
| [15] |
杨颖.日粮能量水平及来源对水貂生产性能和营养物质消化代谢的影响[D].硕士学位论文.北京: 中国农业科学院, 2013: 22-24.
|
| [16] |
LE DIVIDICH J, NOBLET J, BIKAWA T. Effect of environmental temperature and dietary energy concentration on the performance and carcass characteristics of growing-finishing pigs fed to equal rate of gain[J]. Livestock Production Science, 1987, 17: 235-246. DOI:10.1016/0301-6226(87)90069-8 |
| [17] |
NOBLET J, LE DIVIDICH J, BIKAWA T. Interaction between energy level in the diet and environmental temperature on the utilization of energy in growing pigs[J]. Journal of Animal Science, 1985, 61(2): 452-459. |
| [18] |
KORHONEN H, HARRI M, ASIKAINEN J. Thermoregulation of polecat and raccoon dog:a comparative study with stoat, mink and blue fox[J]. Comparative Biochemistry and Physiology Part A:Physiology, 1983, 74(2): 225-230. DOI:10.1016/0300-9629(83)90592-3 |
| [19] |
KORHONEN H, HARRI M, HOHTOLA E. Response to cold in the blue fox and raccoon dog as evaluated by metabolism, heart rate and muscular shivering:a re-evaluation[J]. Comparative Biochemistry and Physiology Part A:Physiology, 1985, 82(4): 959-964. DOI:10.1016/0300-9629(85)90512-2 |
| [20] |
WAMBERG S. Rates of heat and water loss in female mink (Mustela vison) measured by direct calorimetry[J]. Comparative Biochemistry and Physiology Part A:Physiology, 1994, 107(3): 451-458. DOI:10.1016/0300-9629(94)90024-8 |
| [21] |
李昌锐.我国高寒地区水貂日粮蛋白质和能量水平的初步研究[D].硕士学位论文.哈尔滨: 东北林业大学, 2009: 2-5.
|
| [22] |
杨嘉实, 靳世厚, 佟煜人. 貂的能量、蛋白需要及其饲料配制技术研究的综合报告[J]. 特产研究, 1996, 96(4): 1-4. |
| [23] |
于潇滢.环境温度对生长猪生长性能、养分消化及能量代谢的影响[D].硕士学位论文.长春: 吉林农业大学, 2017: 2-8.
|
| [24] |
班志彬, 武斌, 张芳毓, 等. 不同鸡品种维持净能的需要量测定[J]. 动物营养学报, 2020, 32(05): 2158-2163. |
| [25] |
TAUSON A H, ELNIF J, HANSEN N E. Energy metabolism and nutrient oxidation in the pregnant mink (Mustela vison) as a model for other carnivores[J]. The Journal of Nutrition, 1994, 124(Suppl.12): 2609S-26136. |
| [26] |
CHWALIBOG A, JAKOBSEN K, TAUSON A H, et al. Energy metabolism and nutrient oxidation in young pigs and rats during feeding, starvation and re-feeding[J]. Comparative Biochemistry and Physiology Part A:Molecular & Integrative Physiology, 2005, 140(3): 299-307. |
| [27] |
杨嘉实, 冯仰廉. 畜禽能量代谢[M]. 北京: 中国农业出版社, 2004: 30-34.
|
| [28] |
ZHONG W, MU L L, HAN F F, et al. Estimation of the net energy and protein requirements for maintenance of male arctic foxes (Alopex lagopus) during the growth period[J]. Journal of Animal Science, 2019, 97(11): 4579-4587. DOI:10.1093/jas/skz253 |
| [29] |
HARPER R B, TRAVIS H F, GLINSKY M S. Metabolizable energy requirement for maintenance and body composition of growing farm-raised male pastel mink (Mustela vison)[J]. Journal of Nutrition, 1978, 108(12): 1937-1943. DOI:10.1093/jn/108.12.1937 |