




2. 北京首农畜牧发展有限公司, 北京 100076
2. Beijing Sunlon Livestock Development Company Limited, Beijing 100076, China
泌乳早期奶牛对能量的需求快速增加,而采食量却上升缓慢,从而使奶牛能量摄入不足,出现严重的能量负平衡(NEB)[1]。目前的解决方法主要集中在泌乳早期饲喂高代谢蛋白质(MP)和产奶净能(NEL)的饲粮,以提高产奶量,同时避免体况的过度损失及代谢和繁殖问题[2]。近些年的研究倾向于通过添加脂肪提高饲粮能量水平,改善泌乳早期奶牛生产性能和缓解能量负平衡状况[3-4]。膨化全脂大豆(ESB)不仅脂肪含量高(17%~20%),而且是优质的蛋白质饲料(36%~40%),含有大量奶牛必需氨基酸,在提高饲粮能量水平的同时,还可以有效缓解由于能量负平衡而导致的负氮平衡[5]。已有研究表明,在饲粮中添加亚麻籽和ESB,提高了牛奶产量并改变了乳成分[6]。Zhang等[7]研究结果表明在泌乳早期补饲1.5 kg/d ESB可以提高整个泌乳期产奶量。然而,该研究仅设置了1个ESB补饲水平,不一定是最佳饲喂水平。因此,本研究旨在探讨泌乳早期奶牛补饲不同水平的ESB对奶牛生产性能、瘤胃发酵、消化代谢和血液指标的影响,并判断ESB对缓解奶牛能量负平衡的应用效果。
1 材料与方法 1.1 试验动物和饲粮选择4头体重[(587±46) kg]、体况评分(2.95±0.08)和泌乳日龄[(23.6±4.4) d]相近,装有永久性瘤胃瘘管的泌乳早期健康奶牛,采用4×4拉丁方设计,在全混合日粮(TMR)的基础上分别补饲不同水平的ESB:0(对照)、1.0、2.0和3.0 kg/d。ESB购自某饲料厂。饲粮组成及营养水平见表 1。
|
|
表 1 饲粮组成及营养水平(干物质基础) Table 1 Composition and nutrient levels of diets (DM basis) |
试验期从产后第22天开始,共分为4期,每期21 d,其中包括调整适应期14 d,采样期7 d。试验牛为栓系饲养,手工配制TMR,分别于每天的05:30和17:30各饲喂1次,全天自由饮水。记录每天的采食量,剩料量控制在饲喂量的5%~10%。
1.3 样品采集与分析 1.3.1 饲料样品的采集与分析试验期内每周采集1次饲料样品,包括青贮、羊草、苜蓿、ESB、精料混合料、TMR以及剩料,于60 ℃烘箱内烘干48 h制备风干样,粉碎后保存待测。根据AOAC(1990)[8]所描述的方法测定饲料中的干物质(DM)、粗蛋白质(CP)、酸性洗涤纤维(ADF)、粗脂肪(EE)、粗灰分(Ash)、钙(Ca)和磷(P)等常规指标含量,中性洗涤纤维(NDF)含量根据Van Soest等[9]的方法测定。
1.3.2 瘤胃液的采集与分析采样期内每天于采食后第0、4、8、12、16、20和24 h采集瘤胃液,用4层纱布过滤后,立即测定pH;然后再以5 000 r/min的速度离心,收集上清液,并分装于2个塑料瓶中,-20 ℃冷冻保存。其中一瓶根据Broderick等[10]描述的方法,用分光光度计测定氨态氮(NH3-N)含量。另一瓶用25%偏磷酸酸化后,用Agilent 6890N气相色谱仪测定挥发性脂肪酸(VFA)含量[11]。
1.3.3 尿样的收集与分析在每个试验期的采样期内,采用Cao等[12]描述的方法,连续3 d全收粪尿。将每天所收集的粪样混合均匀后,准确称取总粪量的2%,加入1/4粪重的10%浓度的酒石酸,混合均匀并于60 ℃烘干回潮,粉碎后保存待测,粪样中的各种营养成分的分析指标和方法同饲料样品。为防止尿样的腐败分解,在集尿桶中预加入200 mL 10%的稀硫酸以使pH < 3.0,每天取总尿量的2%,用4层纱布过滤后,分装于2个塑料品,于-20 ℃冷冻保存。其中一瓶按照凯氏定氮法测定总氮含量[8],另一瓶采用试剂盒的方法测定尿素氮含量[13]。
1.3.4 血样采集与检测在每个试验期的采样期内于最后1 d晨饲后4 h,使用肝素抗凝一次性真空采血管,通过尾静脉采集血样10 mL,4 000 r/min的速度离心分离血浆,分装于1.5 mL离心管中,于-20 ℃保存待检。血浆中葡萄糖(GLU)、甘油三酯(TG)、总蛋白(TP)、白蛋白(ALB)、总胆红素(TBIL)、非酯化脂肪酸(NEFA)、β-羟基丁酸(BHBA)和尿素氮(UN)含量及谷丙转氨酶(GPT)、谷草转氨酶(GOT)活性使用全自动生化分析仪(Technicon Instruments Corp.,Chauncey,美国)测定;血浆瘦素(Leptin)、胰岛素(Ins)、胰高血糖素(GLN)含量使用放射性免疫试剂盒(Linco Research,Inc.,St. Charles,美国)测定。
1.4 数据统计分析本试验按照拉丁方设计,用SAS 9.1的GLM过程进行分析。瘤胃发酵参数采用以下数学模型:
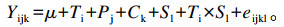
|
式中:Yijk为试验牛在不同饲粮处理下的因变量值;μ为均值;Ti为饲粮处理效应;Pj为试验期效应;Ck为试验牛的随机影响(k=1~4);Sl为采样时间(l=0、4、8、12、16、20、24 h);Ti×Sl为处理与采样时间的互作;eijkl为误差项。
其他数据采用SAS 9.1的GLM方差分析程序进行分析,均值采用Tukey检验进行多重比较。试验数据统计结果以P < 0.05表示差异显著,P < 0.10作为有差异的趋势来讨论。
2 结果与分析 2.1 泌乳早期补饲ESB对奶牛干物质采食量和泌乳性能的影响由表 2可知,泌乳早期补饲ESB有提高奶牛产奶量(P=0.070)和ECM (P=0.068)的趋势。随着ESB补饲水平的增加,乳尿素氮含量显著增加(P < 0.05), 乳脂率有降低的趋势(P=0.081),而非脂乳固体和乳尿素氮含量显著增加(P < 0.05),脂蛋比显著降低(P < 0.05)。
|
|
表 2 泌乳早期补饲不同水平的ESB对奶牛泌乳性能的影响 Table 2 Effects of supplemental different levels of ESB on milk performance of early lactation dairy cows1) |
由表 3可知,随着ESB补饲水平的增加,泌乳早期奶牛瘤胃pH有下降的趋势(P=0.062);NH3-N含量显著增加(P < 0.05);丙酸、戊酸和异戊酸含量显著升高(P < 0.05);ESB补饲水平为2.0 kg/d时,乙酸/丙酸显著低于对照组(P < 0.05)。
|
|
表 3 泌乳早期补饲不同水平的ESB对奶牛瘤胃发酵的影响 Table 3 Effects of supplemental different levels of ESB on ruminal fermentation of early lactation dairy cows |
由表 4可知,随着ESB补饲水平的增加,CP、EE摄入量和粪中EE排出量显著增加(P < 0.05);CP和EE表观消化率显著提高(P < 0.05),DM表观消化率有增加的趋势(P=0.061)。
|
|
表 4 泌乳早期补饲不同水平的ESB对奶牛养分表观消化率的影响 Table 4 Effects of supplemental different levels of ESB on nutrient apparent digestibility of early lactation dairy cows |
由表 5可知,随着ESB补饲水平的增加,总摄入氮、尿氮和尿中尿素氮排泄量、粪尿氮总排泄量、尿氮占总摄入氮的比例以及氮沉积显著增加(P < 0.05),而粪氮占总摄入氮的比例显著下降(P < 0.05);尿素氮占总尿氮的比例有下降的趋势(P=0.063)。
|
|
表 5 泌乳早期补饲不同水平的ESB对奶牛氮代谢的影响 Table 5 Effects of supplemental different levels of ESB on nitrogen metabolism of early lactation dairy cows |
由表 6可知,随着ESB补饲水平的增加,泌乳早期奶牛血浆尿素氮含量显著增加(P < 0.05),而当ESB补饲水平为2.0 kg/d时,血浆总胆红素含量有下降的趋势(P=0.072)。
|
|
表 6 泌乳早期补饲不同水平的ESB对奶牛血液指标的影响 Table 6 Effects of supplemental different levels of ESB on blood parameters in early lactation dairy cows |
本试验中补饲不同水平的ESB对泌乳早期奶牛的干物质采食量没有显著影响,产奶量提高,但乳脂率有下降的趋势,这与前人研究报道结果[14-15]基本一致。Yan等[16]研究表明,在不同玉米青贮、苜蓿青贮条件下,添加或不添加膨化大豆,对干物质采食量、乳蛋白、乳脂和乳糖含量均没有影响。纪鹏等[17]研究表明,在以羊草和玉米青贮为粗饲料的泌乳初期奶牛饲粮中,用膨化大豆替代部分精料使饲粮脂肪含量在5%左右,对干物质采食量没有显著影响,可以提高饲粮养分表观消化率,并且增加产奶量,但是对乳脂的生成有一定程度的抑制作用。Kennelly等[18]和Dhiman等[19]的研究结果表明,当饲粮脂肪含量小于6%时,添加脂肪的种类和数量对泌乳牛干物质采食量都不会有显著影响[20]。但也有研究表明,添加油脂对奶牛干物质采食量有一定的抑制作用,其抑制程度可能与油脂添加量和饲粮结构有关[21-22]。本试验中,补饲ESB的奶牛干物质采食量更高,可能是因为补饲ESB导致能量和蛋白质摄入量增加[23]。
在各组干物质采食量无显著差异的情况下,补饲ESB组产奶量有提高的趋势。一方面可能是因为ESB增加了饲粮中脂肪含量,提高了饲粮能量水平及养分的消化率,从而促进了泌乳,这与Meignan等[24]研究结果一致。另一方面,膨化处理后抑制了大豆中抗营养因子活性并增加了大豆过瘤胃蛋白的比例,从而为小肠段消化提供了更多的优质蛋白质和必需氨基酸,促进了泌乳潜力的发挥[25]。试验组较高的乳蛋白、乳糖和SNF产量则可能是由于较高的产奶量和饲粮养分消化率所致。本试验中,补饲ESB后乳脂率有降低的趋势,与Zhang等[7]的报道结果一致。这一方面可能与较多的产奶量对乳成分的稀释作用有关,另一方面可能与补饲ESB后瘤胃中乙酸/丙酸降低有关。乙酸是合成乳脂重要的前体物,乙酸和丙酸之间存在适当的比例。因为乙酸合成脂肪所需的还原型烟酰胺腺嘌呤二核苷酸磷酸(NADPH)主要是由GLU经磷酸戊糖途径生成,当细胞质中NADPH的含量不足时,会限制乙酸的利用;而丙酸是反刍动物GLU异生的主要前体物质,所以只有乙酸/丙酸适当,才能保证乙酸的充分利用,进而保证一定的乳脂率。Dawod等[6]报道,泌乳牛饲粮中添加膨化亚麻籽和ESB可以提高产奶量,但降低乳脂率。这可能是因为在乳腺细胞中,外源性的脂肪酸与从头合成的短链脂肪酸在脂化过程中发生相互竞争,外源性的脂肪酸可对产脂酶发生负反馈作用,抑制了脂肪酸的从头合成,从而降低了乳脂中短链脂肪酸的含量。另外,还有大量研究表明,饲粮添加未保护脂肪会在一定程度上抑制微生物尤其是纤维分解菌的活性,进而降低纤维消化,减少乙酸生成,从而降低乳脂率。膨化处理使含脂量很高的大豆中脂肪暴露出来,这有可能是导致试验组奶牛乳脂率降低的主要原因。除脂肪因素外,ESB中较高的蛋白质含量也可能是产奶量和乳成分的重要影响因子。Depeters等[26]认为,饲粮CP水平对奶牛的产奶量以及乳成分的作用是通过影响CP摄入量、CP与饲粮能量间的互作实现的。
另外,本研究还发现,随着ESB补饲水平的增加,泌乳早期奶牛乳尿素氮含量显著增加,这可能是由于ESB中蛋白质含量较高,补饲奶牛的氮摄入量明显增加造成的。与Giallongo等[27]结果一致。Dawod等[6]研究表明,在基础饲粮基础上,按13 ∶ 3的比例添加膨化亚麻籽和膨化大豆可以显著提高乳尿素氮和血浆尿素氮含量。Harper等[14]用ESB替代溶剂提取豆粕饲喂泌乳奶牛发现对乳尿素氮含量没有显著影响。Webb等[28]在评价棉籽粕完全替代豆粕饲喂泌乳奶牛的试验中发现,饲喂棉籽粕的奶牛乳尿素氮含量降低,即豆粕使乳尿素氮含量增加。前人在统计大量的研究结果后发现,饲粮CP水平与乳尿素氮含量显著相关,饲粮CP水平是影响乳尿素氮含量程度最大的营养因素。Nousiainen等[29]认为乳尿素氮含量与摄入氮和饲粮CP水平高度相关,乳尿素氮含量可以用来预测奶牛饲粮CP水平。
3.2 泌乳早期补饲ESB对奶牛瘤胃发酵的影响大量试验结果表明,饲粮中添加脂肪对瘤胃的pH变化影响不大,但对挥发性脂肪酸含量和比例、NH3-N的含量及微生物的组成和比例有不同程度的影响[30-31]。本研究发现,随着ESB补饲水平的增加,泌乳早期奶牛瘤胃pH有下降的趋势,NH3-N含量显著增加,丙酸、戊酸和异戊酸含量显著升高,当ESB补饲水平为2.0 kg/d时,乙酸/丙酸显著低于对照组。ESB中除含有较多脂肪外,还含有丰富的蛋白质和部分碳水化合物,这可能是与直接添加脂肪对瘤胃发酵产生影响的主要原因。Van Houtert[32]认为,瘤胃微生物达到最大生长速度的适宜pH在5.7以上。因为,瘤胃pH决定着瘤胃微生物对底物的利用效率。当瘤胃pH <6.2时微生物合成蛋白质的效率下降[25]。本试验中,当ESB补饲水平为2.0和3.0 kg/d时,瘤胃平均pH低于6.2,表明在此添加水平下,瘤胃微生物蛋白合成效率降低。
瘤胃NH3-N主要来源于食糜中的蛋白质和非蛋白氮(NPN)的降解,瘤胃NH3-N含量的高低在一定程度上反映了蛋白质降解与合成间所达到的平衡状况。瘤胃内NH3-N作为纤维分解菌的必需氮源、非纤维分解菌的部分氮源,是影响微生物活力的一项重要指标。关于微生物最佳生长的NH3-N含量,Satter等[33]试验结果表明,NH3-N含量小于5 mg/dL时发酵的“解偶联”作用引起微生物的产量降低,生产效率下降,该值被广泛用作瘤胃最低NH3-N含量的标准。Preston等[34]报道微生物生长对NH3-N含量耐受的临界范围为6~30 mg/dL。本试验中各组奶牛瘤胃NH3-N含量均在适宜范围之内,但是随着ESB补饲水平的增加,瘤胃NH3-N含量显著增加,这可能是因为ESB中的瘤胃降解蛋白质含量较高造成的。因此,如果ESB补饲水平过大,有可能会导致瘤胃能氮不平衡,微生物无法有效利用合成微生物蛋白而造成氮损失。
反刍动物的消化能60%~80%由瘤胃挥发性脂肪酸提供,挥发性脂肪酸为奶牛的代谢过程和牛奶合成提供了大部分的能量前体[35],同时,挥发性脂肪酸也是奶牛乳成分的重要合成原料,其中乙酸是乳脂合成前体,而丁酸对乳脂合成也有正效应,丙酸则有利于乳糖的合成且降低乳脂率。Woodford等[36]研究表明,乙酸/丙酸大于2.5时可维持正常的乳脂合成。本试验中各组奶牛瘤胃中乙酸/丙酸在3.0~3.6,因此均能维持奶牛正常合成乳脂的需要。但是随着ESB补饲水平的增加,丙酸含量显著升高,ESB补饲水平为2.0 kg/d时,乙酸/丙酸显著低于对照组。这可能是因为植物油中的不饱和脂肪酸对瘤胃微生物所产生的毒害作用,对瘤胃的发酵产生了一定的负面影响[37]。有研究表明,大豆油籽中的多不饱和脂肪酸影响瘤胃发酵模型,降低了乙酸/丙酸[13, 38]。
3.3 泌乳早期补饲ESB对奶牛养分表观消化率和氮代谢的影响很多研究结果表明,当添加过多的油脂或者改变饲粮中的油脂比例,可能会对瘤胃微生物产生毒害作用,进而影响营养物质的消化率[39-40]。本试验中随着ESB补饲水平的增加,NDF和ADF表观消化率虽然有所降低,但差异不显著。推测可能是因为油脂在瘤胃中迅速释放,从而抑制瘤胃微生物对纤维的降解[41]。但Harper等[14]在比较奶牛对ESB和溶剂提取豆粕(SSB)消化率发现,即使ESB的EE含量比SSB高23%~25%,对全肠道消化率并没有影响。本研究还发现,随着ESB补饲水平的增加,CP和EE表观消化率显著增加,DM表观消化率有增加的趋势,与纪鹏等[17]的研究结果一致。但胡志勇等[42]发现,在高粗料奶牛饲粮(精粗比为40 ∶ 60)中,添加4%豆油、4%胡麻油及2%豆油+2%胡麻油后,对奶牛干物质采食量、瘤胃发酵、有机物质和纤维物质瘤胃及其全肠道消化率没有显著影响。Giallongo等[27]研究结果表明,饲粮添加ESB与SSB相比,对所有养分的消化率增加或有增加的趋势。
氮代谢是动物氮需要量的研究基础,深入了解动物的氮代谢有助于保证满足动物氮的营养需要,在发挥最大生产性能的同时又能尽量减少氮的排泄量。在本试验中,尿氮排出量随氮的摄入量增加而显著增加,而粪氮排出量比较稳定。Tamminga等[43]指出,年单产为6 250 kg鲜乳的荷斯坦奶牛,氮的采食量在175 kg/年,仅有20%的氮转化为牛奶或沉积到体组织,而28.5%以粪氮形式排出,50.3%以尿氮形式排出。有研究表明,增加饲粮CP摄入量能够导致尿氮排泄量迅速增加[44],超过动物蛋白质需要量的几乎所有的氮都是通过尿液排出体外[45]。因此,粪氮排泄量相当稳定,与干物质采食量呈一定比例。大多数奶牛饲粮氮消化率很高,微生物蛋白消化率也很高,提高奶牛氮消化率可能性不大[44]。这可能是本试验和以往研究中粪氮排泄量未显著受到饲粮CP水平影响的主要原因。Mulligan等[46]的研究表明,奶牛氮摄入量与粪氮、尿氮及总氮排泄量呈正相关关系。本试验中,随着ESB补饲水平的增加,也发现了相似的结果,这可能是因为ESB中CP含量较高,补饲ESB后奶牛摄入氮显著增加,被动物机体消化吸收后不被利用的部分主要以尿素的形式排放于尿液中。但也有研究表明奶牛粪、尿氮排泄量不随氮摄入量的增加而增加[14, 26]。
多数学者认为当奶牛饲粮中CP水平在一定水平时,产奶量和乳蛋白产量最大,而奶牛氮的排泄量相对较低,如果CP水平超过这个值,产奶量下降或者没有影响,说明饲粮CP水平过高。本试验中当ESB补饲水平达到2.0 kg/d以上时,产奶量增幅很小,表明在本试验条件下,当饲粮CP达到这个水平时(17.13%)已满足或超过奶牛泌乳和维持需要。Groff等[47]的研究发现,对于35.5~40.0 kg/d产奶量的奶牛来说,饲粮15%~17% CP水平对维持产奶有益,超过17%后对生产性能无益。另外,由于受反刍动物瘤胃影响,奶牛氮代谢过程不易量化,但通过对乳尿素氮和血浆尿素氮含量的检测,可以从侧面了解奶牛机体氮代谢状况,从而评估奶牛的蛋白质营养状况和饲粮中CP与能量摄入平衡情况。Carder等[48]也认为,泌乳早期补充MP或氨基酸对生产性能产生的影响可以是短期的(产奶量),也可以是长期的(乳成分)。
3.4 泌乳早期补饲ESB对奶牛血液指标的影响血浆尿素氮大部分是氨基酸在肝脏代谢而产生或从大肠吸收的氨合成的,因此血浆尿素氮含量可反映动物蛋白质代谢状况,作为机体蛋白质沉积的一个指标。本研究中随着ESB补饲水平的增加,泌乳早期奶牛血浆尿素氮含量显著增加。这可能主要是由于补饲ESB后,饲粮CP水平显著增加,奶牛摄入氮增加造成的。有研究表明,血浆尿素氮与蛋白质摄入量呈正相关[49],血浆中尿素含量高主要来源于饲粮中高含量的蛋白质降解产物。根据Whitelaw等[50]的研究,血浆尿素氮与瘤胃NH3-N含量存在线性相关。因此本试验中补饲ESB后,奶牛瘤胃NH3-N含量显著增加,也是血浆尿素氮含量增加的重要原因。另外,由于尿素是小分子物质,血液中的尿素氮通过向乳中扩散,随着牛奶排出体外,同时乳和血液中的尿素氮含量也保持着一定的动态平衡,二者紧密正相关。Baker等[51]研究表明,血浆尿素氮含量具有在1 d中的变化幅度较大的特点,不利于不同试验结果的比较,其在奶牛营养中的应用受到了很大的限制,而乳尿素氮含量能代表 1 d中血浆尿氮含量的平均水平,因此乳尿素氮含量可以更准确地反映机体氮代谢的情况。再加上测定乳尿素氮含量的取样简单方便,测定容易、准确,这使得乳尿素氮含量测定在奶牛生产中的应用越来越广泛。
胆红素是血液循环中衰老红细胞在肝脏、脾脏及骨髓的单核-吞噬细胞系统中分解和破坏的产物。血液中的胆红素有80%~85%来源于红细胞中血红蛋白的分解,其余15%~20%来自骨髓内红细胞前体细胞的破坏及其他组织中的血红素蛋白,进入血液中的胆红素与白蛋白结合后输送到肝脏代谢,经肝细胞转化后排入毛细胆管并随胆汁排出,若机体内生成胆红素过多超过了肝细胞的处理能力,或者肝细胞处理胆红素的能力下降,以及胆红素排泄障碍等均可使血中胆红素含量增高。胆红素含量是反映肝功能的指标之一[52]。如果肝细胞发生脂变性,血中的胆红素就不易进入肝脏,这样形成的大分子物质就很难进入肝细胞。血中胆红素含量升高幅度不明显时,其对肝细胞损害就不是一个灵敏的指标,但肝脏疾病中胆红素含量明显增高,常反映出肝细胞有严重的损害。本试验中,当ESB补饲水平为2.0 kg/d时,血浆总胆红素含量有下降的趋势,与Zhang等[7]研究结果一致。这表明泌乳早期奶牛补饲ESB后改善了能量平衡,减轻了肝脏负荷,从而引起血浆总胆红素含量的降低。
4 结论泌乳早期奶牛补饲ESB后,CP、EE和DM的表观消化率以及氮沉积量增加,但乳脂率有下降趋势。当ESB补饲水平为2.0 kg/d时,血浆总胆红素含量有下降的趋势;当补饲水平为2.0和3.0 kg/d时,瘤胃液丙酸含量显著升高,可促进肝糖原异生,增加泌乳早期奶牛的能量供给,但可能会导致瘤胃微生物蛋白合成效率降低。随ESB的补饲水平增加,尿氮排出量增加,而粪氮排出量比较稳定,同时乳尿素氮和血浆尿素氮含量显著提高。综合考虑,在本试验条件下,泌乳早期奶牛补饲1.0~2.0 kg/d ESB,有利于缓解能量负平衡并提高生产性能。
| [1] |
DRACKLEY J K. Biology of dairy cows during the transition period: the final frontier?[J]. Journal of Dairy Science, 1999, 82(11): 2259-2273. DOI:10.3168/jds.S0022-0302(99)75474-3 |
| [2] |
DANN H M. Nutritional management strategies for dry and fresh cows. Large dairy herd management[M]. 3rd.ed. Champaign, IL: American Dairy Science Association, 2017: 699-712.
|
| [3] |
ZHANG Z, LA S K, ZHANG G W, et al. Diet supplementation of palm fat powder and coated folic acid on performance, energy balance, nutrient digestion, ruminal fermentation and blood metabolites of early lactation dairy cows[J]. Animal Feed Science and Technology, 2020, 265: 114520. DOI:10.1016/j.anifeedsci.2020.114520 |
| [4] |
王雪, 秦贵信, 甄玉国, 等. 日粮碳水化合物与脂肪对奶牛采食量和能量分配影响的研究进展[J]. 中国畜牧杂志, 2018, 54(5): 11-14. WANG X, QIN G X, ZHEN Y G, et al. Research progress on the effects of different dietary energy sources on feed intake and energy partition in dairy cows[J]. Chinese Journal of Animal Science, 2018, 54(5): 11-14 (in Chinese). |
| [5] |
蒋林树, 孟庆翔, 马俊云, 等. 膨化全脂大豆对高产奶牛乳脂中PUFA含量的影响[J]. 中国农学通报, 2007, 23(2): 5-9. JIANG L S, MENG Q X, MA J Y, et al. Study on the effects of feeding EFFSB on the content of PUFA of milk fat of milking cows[J]. Chinese Agricultural Science Bulletin, 2007, 23(2): 5-9 (in Chinese). DOI:10.3969/j.issn.1000-6850.2007.02.002 |
| [6] |
DAWOD A, AHMED H, ABOU-ELKHAIR R, et al. Effects of extruded linseed and soybean dietary supplementation on lactation performance, first-service conception rate, and mastitis incidence in Holstein dairy cows[J]. Animals, 2020, 10(3): 436. DOI:10.3390/ani10030436 |
| [7] |
ZHANG Q, SU H W, WANG F W, et al. Effects of energy density in close-up diets and postpartum supplementation of extruded full-fat soybean on lactation performance and metabolic and hormonal status of dairy cows[J]. Journal of Dairy Science, 2015, 98(10): 7115-7130. DOI:10.3168/jds.2014-9112 |
| [8] |
A.O.A.C. Official methods of analysis[M]. 15th ed. Washington, D.C.: Association of Official Analytical Chemist, 1990.
|
| [9] |
VAN SOEST P J, ROBERTSON J B, LEWIS B A. Methods for dietary fiber, neutral detergent fiber, and nonstarch polysaccharides in relation to animal nutrition[J]. Journal of Dairy Science, 1991, 74(10): 3583-3597. DOI:10.3168/jds.S0022-0302(91)78551-2 |
| [10] |
BRODERICK G A, KANG J H. Automated simultaneous determination of ammonia and total amino acids in ruminal fluid and in vitro media[J]. Journal of Dairy Science, 1980, 63(1): 64-75. DOI:10.3168/jds.S0022-0302(80)82888-8 |
| [11] |
ERWIN E S, MARCOAND G J, EMERY E M. Volatile fatty acid analysis of food and rumen fluid by gas chromatography[J]. Journal of Dairy Science, 1961, 44(9): 1768-1777. DOI:10.3168/jds.S0022-0302(61)89956-6 |
| [12] |
CAO Z J, MA M, YAN X Y, et al. A simple urine-collecting apparatus and method for cows and heifers[J]. Journal of Dairy Science, 2009, 92(10): 5224-5228. DOI:10.3168/jds.2009-2036 |
| [13] |
BRODERICK G A, CLAYTON M K. A statistical evaluation of animal and nutritional factors influencing concentrations of milk urea nitrogen[J]. Journal of Dairy Science, 1997, 80(11): 2964-2971. DOI:10.3168/jds.S0022-0302(97)76262-3 |
| [14] |
HARPER M T, OH J, MELGAR A, et al. Production effects of feeding extruded soybean meal to early-lactation dairy cows[J]. Journal of Dairy Science, 2019, 102(10): 8999-9016. DOI:10.3168/jds.2019-16551 |
| [15] |
AMANLOU H, MAHERI-SIS N, BASSIRI S, et al. Nutritional value of raw soybeans, extruded soybeans, roasted soybeans and tallow as fat sources in early lactating dairy cows[J]. Open Veterinary Journal, 2012, 2(1): 88-94. |
| [16] |
YAN R, HAN J G, ZHANG X, et al. Effects of different corn silage: alfalfa silage ratios and full fat extruded soybeans on milk composition, conjugated linoleic acids content in milk fat and performance of dairy cows[J]. African Journal of Biotechnology, 2011, 9(33): 5464-5465. |
| [17] |
纪鹏, 陈萍, 李胜利, 等. 不同油料籽实日粮对奶牛养分表观消化率、生产性能及血液指标的影响[J]. 动物营养学报, 2008, 20(2): 217-222. JI P, CHEN P, LI S L, et al. Effects of oilseeds on apparent digestibility, milking performance and blood indices of dairy cows in early lactation[J]. Chinese Journal of Animal Nutrition, 2008, 20(2): 217-222 (in Chinese). DOI:10.3969/j.issn.1006-267X.2008.02.017 |
| [18] |
KENNELLY J J. The fatty acid composition of milk fat as influenced by feeding oilseeds[J]. Animal Feed Science and Technology, 1996, 60(3/4): 137-152. |
| [19] |
DHIMAN T R, SATTER L D, PARIZA M W, et al. Conjugated linoleic acid (CLA) content of milk from cows offered diets rich in linoleic and linolenic acid[J]. Journal of Dairy Science, 2000, 83(5): 1016-1027. DOI:10.3168/jds.S0022-0302(00)74966-6 |
| [20] |
PETIT H V. Review: feed intake, milk production and milk composition of dairy cows fed flaxseed[J]. Canadian Journal of Animal Sciencei, 2010, 90(2): 115-127. DOI:10.4141/CJAS09040 |
| [21] |
MARTIN C, ROUEL J, JOUANY J P, et al. Methane output and diet digestibility in response to feeding dairy cows crude linseed, extruded linseed, or linseed oil[J]. Journal of Animal Science, 2008, 86(10): 2642-2650. DOI:10.2527/jas.2007-0774 |
| [22] |
LERCH S, FERLAY A, POMIōS D, et al. Rapeseed or linseed supplements in grass-based diets: effects on dairy performance of Holstein cows over 2 consecutive lactations[J]. Journal of Dairy Science, 2012, 95(4): 1956-1970. DOI:10.3168/jds.2011-4575 |
| [23] |
SOLOMON R, CHASE L E, BEN-GHEDALIA D, et al. The effect of nonstructural carbohydrate and addition of full fat extruded soybeans on the concentration of conjugated linoleic acid in the milk fat of dairy cows[J]. Journal of Dairy Science, 2000, 83(6): 1322-1329. DOI:10.3168/jds.S0022-0302(00)74998-8 |
| [24] |
MEIGNAN T, LECHARTIER C, CHESNEAU G, et al. Effects of feeding extruded linseed on production performance and milk fatty acid profile in dairy cows: a Meta-analysis[J]. Journal of Dairy Science, 2017, 100(6): 4394-4408. DOI:10.3168/jds.2016-11850 |
| [25] |
Subcommittee on Dairy Cattle Nutrition, Committee on Animal Nutrition, Board on Agriculture and Natural Resources, et al. Nutrient requirements of dairy cattle[M]. 7th ed. Washington, D.C: National Academy Press, 2001.
|
| [26] |
DEPETERS E J, CANT J P. Nutritional factors influencing the nitrogen composition of bovine milk: a review[J]. Journal of Dairy Science, 1992, 75(8): 2043-2070. DOI:10.3168/jds.S0022-0302(92)77964-8 |
| [27] |
GIALLONGO F, OH J, FREDERICK T, et al. Extruded soybean meal increased feed intake and milk production in dairy cows[J]. Journal of Dairy Science, 2015, 98(9): 6471-6485. DOI:10.3168/jds.2015-9786 |
| [28] |
WEBB N W, BERNARD PAS J K, TAO S. Production responses to diets supplemented with soybean meal, expeller soybean meal, or dry-extruded cottonseed cake by lactating dairy cows[J]. Applied Animal Science, 2019, 35(6): 543-549. DOI:10.15232/aas.2019-01906 |
| [29] |
NOUSIAINEN J, SHINGFIELD K J, HUHTANEN P. Evaluation of milk urea Nitrogen as a diagnostic of protein feeding[J]. Journal of Dairy Science, 2004, 87(2): 386-398. DOI:10.3168/jds.S0022-0302(04)73178-1 |
| [30] |
ZEIDALI-NEJAD A, GHORBANI G R, KARGAR S, et al. Nutrient intake, rumen fermentation and growth performance of dairy calves fed extruded full-fat soybean as a replacement for soybean meal[J]. Animal, 2018, 12(4): 733-740. DOI:10.1017/S1751731117002154 |
| [31] |
PALMQUIST D L, GRIINARI J M. Milk fatty acid composition in response to reciprocal combinations of sunflower and fish oils in the diet[J]. Animal Feed Science and Technology, 2006, 131(3/4): 358-369. |
| [32] |
VAN HOUTERT M F J. The production and metabolism of volatile fatty acids by ruminants fed roughages: a review[J]. Animal Feed Science and Technology, 1993, 43(3/4): 189-225. |
| [33] |
SATTER L D, SLYTER L L. Effect of ammonia concentration of rumen microbial protein production in vitro[J]. The British Journal of Nutrition, 1974, 32(2): 199-208. DOI:10.1079/BJN19740073 |
| [34] |
PRESTON H, LENG D. Automated simultaneous determination of ammonia and total amino acid in ruminal fluids and in vitro media[J]. Journal of Animal Science, 1987, 63: 64-67. |
| [35] |
HUANG W M, TIAN Y J, LI S L, et al. Reduced energy density of close-up diets decrease ruminal pH and increase concentration of volatile fatty acids postpartum in Holstein cows[J]. Animal Science Journal, 2017, 88(11): 1700-1708. DOI:10.1111/asj.12835 |
| [36] |
WOODFORD S T, MURPHY M R. Effect of forage physical form on chewing activity, dry matter intake, and rumen function of dairy cows in early lactation[J]. Journal of Dairy Science, 1988, 71(3): 674-686. DOI:10.3168/jds.S0022-0302(88)79606-X |
| [37] |
BROUDISCOU L, POCHET S, PONCET C. Effect of linseed oil supplementation on feed degradation and microbial synthesis in the rumen of ciliate-free and refaunated sheep[J]. Animal Feed Science and Technology, 1994, 49(3/4): 189-202. |
| [38] |
高军肖. 植物油(籽)对牛奶共扼亚油酸前体物累积规律影响的研究[D]. 硕士学位论文. 北京: 中国农业科学院, 2004. GAO J X. The effects of plant oil and oilseed an accumulation of precursors of milk conjugated linoleic acid[D]. Master's Thesis. Beijing: Chinese Academy of Agricultural Sciences, 2004. (in Chinese) |
| [39] |
DE SOUZA J, PRESEAULT C L, LOCK A L. Altering the ratio of dietary palmitic, stearic, and oleic acids in diets with or without whole cottonseed affects nutrient digestibility, energy partitioning, and production responses of dairy cows[J]. Journal of Dairy Science, 2018, 101(1): 172-185. DOI:10.3168/jds.2017-13460 |
| [40] |
DE SOUZA J, PROM C M, LOCK A L. Altering the ratio of dietary palmitic and oleic acids affects nutrient digestibility, metabolism, and energy balance during the immediate postpartum in dairy cows[J]. Journal of Dairy Science, 2021, 104(3): 2910-2923. DOI:10.3168/jds.2020-19312 |
| [41] |
PALMQUIST D L, JENKINS T C. Fat in lactation rations: review[J]. Journal of Dairy Science, 1980, 63(1): 1-14. DOI:10.3168/jds.S0022-0302(80)82881-5 |
| [42] |
胡志勇, 王加启, 卜登攀, 等. 奶牛日粮添加豆油和胡麻油对有机物和纤维物质消化率的影响[J]. 中国畜牧兽医, 2007, 34(6): 5-8. HU Z Y, WANG J Q, BU D P, et al. Effect of supplementation of soybean oil and linseed oil on digestibility of organic matters and fibers in dairy cows[J]. China Animal Husbandry & Veterinary Medicine, 2007, 34(6): 5-8 (in Chinese). DOI:10.3969/j.issn.1671-7236.2007.06.001 |
| [43] |
TAMMINGA S, VAN STRAALEN W M, SUBNEL A P J, et al. The Dutch protein evaluation system: the DVE/OEB-system[J]. Livestock Production Science, 1994, 40(2): 139-155. DOI:10.1016/0301-6226(94)90043-4 |
| [44] |
TAMMINGA S. Nutrition management of dairy cows as a contribution to pollution control[J]. Journal of Dairy Science, 1992, 75(1): 345-357. DOI:10.3168/jds.S0022-0302(92)77770-4 |
| [45] |
PEYRAUD J L, VERITÉAND R, DELABY L. Nitrogen excretion by dairy cows: effect of the diet and of the level of production[J]. Fourrages, 1995, 142: 131-144. |
| [46] |
MULLIGAN F J, DILLON P, CALLAN J J, et al. Supplementary concentrate type affects nitrogen excretion of grazing dairy cows[J]. Journal of Dairy Science, 2004, 87(10): 3451-3460. DOI:10.3168/jds.S0022-0302(04)73480-3 |
| [47] |
GROFF E B, WU Z. Milk production and Nitrogen excretion of dairy cows fed different amounts of protein and varying proportions of alfalfa and corn silage[J]. Journal of Dairy Science, 2005, 88(10): 3619-3632. DOI:10.3168/jds.S0022-0302(05)73047-2 |
| [48] |
CARDER E G, WEISS W P. Short- and longer-term effects of feeding increased metabolizable protein with or without an altered amino acid profile to dairy cows immediately postpartum[J]. Journal of Dairy Science, 2017, 100(6): 4528-4538. DOI:10.3168/jds.2016-12362 |
| [49] |
HOF G, VERVOORN M D, LENAERS P J, et al. Milk urea nitrogen as a tool to monitor the protein nutrition of dairy cows[J]. Journal of Dairy Science, 1997, 80(12): 3333-3340. |
| [50] |
WHITELAW F G, MILNE J S, CHEN X B. The effect of a rumen microbial fermentation on urea and nitrogen metabolism of sheep nourished by intragastric infusion[J]. Experimental Physiology, 1991, 76(1): 91-101. DOI:10.1113/expphysiol.1991.sp003484 |
| [51] |
BAKER L D, FERGUSON J D, RAMBERG C F. Kinetic analysis of urea transport from plasma to milk in dairy cows[J]. Journal of Dairy Science, 1992, 75(Suppl.1): 181. |
| [52] |
GRAUGNARD D E, BIONAZ M, TREVISI E, et al. Blood immunometabolic indices and polymorphonuclear neutrophil function in peripartum dairy cows are altered by level of dietary energy prepartum[J]. Journal of Dairy Science, 2012, 95(4): 1749-1758. DOI:10.3168/jds.2011-4579 |