




2. 新希望六和股份有限公司, 北京 100102
2. New Hope Liuhe Co., Ltd., Beijing 100102, China
在实际生产中饲料有效能的成本占肉鸭饲粮配方成本的60%以上,因此,准确地获得肉鸭对常用饲料原料的代谢能(ME)数据非常重要。而通过试验饲粮的ME计算待测饲料原料的ME时,计算方式将直接影响ME的数据准确性。因此,选择合适的计算方式是确保ME准确性的基本前提。在《肉鸭饲养标准》(NY/T 2122-2012)关于鸭饲料表观代谢能(AME)测定方法的描述中,AME的计算方法仅适合于直接测定的饲粮或饲料原料,而对于必须采用套算法测定的蛋白质饲料原料却未作阐述。传统上,套算法测定饲料原料的ME都是根据试验饲粮中基础饲粮和待测饲料原料的比例及试验饲粮和基础饲粮的ME计算得出待测饲料原料的ME[1]。然而,该方法必须以待测饲料原料和试验饲粮的总能可加为基本前提,但实际生产中由于待测饲料原料和基础饲粮间的加和效应以及氧弹热量计的内在变异导致总能的计算值和实测值的差值最高可达270 kJ/kg DM[2],由此计算出的酶水解物能值与实测值的差值在210 kJ/kg DM以上[3]。Adeola[4]、Woyengo等[5]、Kong等[6]在猪的试验中根据试验饲粮中基础饲粮和待测饲料原料的比例及试验饲粮和基础饲粮的能量消化(代谢)率计算得出待测饲料原料的能量消化(代谢)率,然后通过能量消化(代谢)率乘以总能获得待测饲料原料的有效能值。类似地,尹玉港等[7]比较了通过肉鸭试验饲粮和基础饲粮的ME计算待测脂肪的ME与通过脂肪消化率和脂肪的总能值计算的脂肪的ME,得出了消化率计算法的ME变异度更低。由此可见,在套算法中对同一代谢试验数据采用不同的计算方式得出的待测饲料原料的ME有所差异。为此,本研究探讨套算法和消化率计算法这2种计算方式得出的肉鸭饲料原料ME的差异及对饲粮中饲料原料间ME可加性的影响,以期为肉鸭饲料原料ME的测定中选择合适的计算方式提供参考。
1 材料与方法 1.1 试验设计本研究分为2个试验。试验一:首先采用绝食法分5个批次测定肉鸭内源能损失的变异,然后通过排空强饲法测定10个肉鸭饲料原料的ME,其中玉米、小麦、高粱、大麦采用直接法测定,豆粕、棉籽粕、米糠、菜籽饼、玉米干酒糟及其可溶物(DDGS)、小麦麸采用差量法测定。通过2种公式计算ME,公式1为套算法,是根据基础饲粮、试验饲粮的ME及试验饲粮中基础饲粮的比例计算待测饲料原料的ME;公式2为消化率计算法,是根据待测饲料原料的能量消化率乘以待测饲料原料的总能得出ME,其中待测饲料原料的能量消化率根据基础饲粮、试验饲粮的能量消化率及试验饲粮中基础饲粮和待测原料的供能比例计算。试验二:以试验一中的10个肉鸭饲料原料配制10种饲粮,采用排空强饲法测定其ME,同时根据饲粮中各原料的组成比例及试验一中2种计算方式得出的原料ME计算饲粮的ME,比较饲粮ME计算值与实测值的差异。
1.2 饲料原料及试验饲粮试验一中的10个饲料原料--玉米、小麦、高粱、大麦、大豆粕、棉籽粕、米糠、菜籽饼、玉米DDGS、小麦麸由新希望六和股份有限公司提供,其营养水平见表 1。
|
|
表 1 试验饲料原料的营养水平(风干基础) Table 1 Nutrient levels of test feed ingredients (air-dry basis) |
在ME测定中,试验饲粮的配制概述如下:4种谷实类原料玉米、小麦、高粱、大麦配制的试验饲粮(饲粮1~4)由待测饲料原料加磷酸氢钙、石粉、氯化钠、维生素-微量元素预混料组成,6种蛋白质类和其他类原料大豆粕、棉籽粕、米糠、菜籽饼、玉米DDGS、小麦麸配制的试验饲粮(饲粮5~10)由玉米、待测饲料原料加磷酸氢钙、石粉、氯化钠、维生素-微量元素预混料组成,使其蛋白质水平保持在20%左右(表 2)。
|
|
表 2 试验饲粮组成(风干基础,试验一) Table 2 Composition of experimental diets (air-dry basis, experiment 1) |
试验二的10种验证饲粮中饲粮1~3由玉米、豆粕、小麦、小麦麸中的2种或2种以上与磷酸氢钙、石粉、氯化钠、维生素-微量元素预混料组成,饲粮1中原料的供能比值与饲粮2~3是一致的;饲粮4~8由玉米、玉米DDGS、米糠、棉籽粕、菜籽饼、小麦麸中的2种或2种以上原料与磷酸氢钙、石粉、氯化钠、维生素-微量元素预混料组成,饲粮4中原料的供能比值与饲粮5~8是一致的;饲粮9~10由小麦、玉米、高粱、大麦中的3种或3种以上原料磷酸氢钙、石粉、氯化钠、维生素-微量元素预混料组成,饲粮9中原料的供能比值与饲粮10是一致的(表 3)。
|
|
表 3 验证饲粮组成(风干基础,试验二) Table 3 Composition of the validation diets (air-dry basis, experiment 2) |
上述每个试验饲粮制备4~5 kg用于代谢试验。在制备过程中,所有原料按各试验饲粮重量要求分别称量,并放入小塑料桶中,混合后粉碎过1 mm筛。粉碎好的试验饲粮经多次手工混合后,加入适量的水,冷制粒成直径3 mm、长度6 mm的颗粒料,待试验饲粮风干至干物质含量在88%以上时,置于自封袋中保存备用。
1.3 肉鸭试验饲粮ME的测定每个试验饲粮ME或内源排泄量的测定选择健康、体重(平均体重约为3.7 kg)基本一致的成年樱桃谷公鸭12只,随机分为4个重复,每个重复3只鸭。在代谢试验中,适应期试验鸭的饲养管理按动物营养学国家重点实验室常规程序进行,自然光照。
肉鸭饲料ME测定过程参照Sibbald[8]和NY/T 2122-2012的方法进行,具体过程为:适应期72 h,饲喂商品饲粮(新希望六和股份有限公司生产);预试期24 h,饲喂试验饲粮;禁饲排空36 h后强饲试验饲粮60 g,排泄物收集36 h。适应期前,将试验鸭肛门周围的羽毛拔净,将直径4 cm的瓶盖缝合到肛门周边的皮脂上,保证盖子与肛门周围的皮肤接触紧密。准确称取60 g用于强饲的试验饲粮,同步测定饲粮的干物质含量。强饲过程中分别记录每只鸭的强饲结束时间。排泄物的收集参照Adeola等[9]设计的方案进行,每次待集粪袋中的排泄物超过粪袋容积的1/4时,将排泄物无损失地转入相应编号的培养皿中,待对每只试验鸭准确收集了36 h的粪便后,将全部粪样转入65 ℃烘箱中鼓风干燥72 h。烘干样在室温回潮24 h后称重,然后立即粉碎过40目筛制成风干样品,并测定粪便样品的干物质含量。
1.4 化学分析饲料、粪样的总能用全自动氧弹计(型号PARR-6400;以苯甲酸为标准物进行校准)进行测定,水分、粗蛋白质、粗脂肪、粗灰分、粗纤维、中性洗涤纤维、酸性洗涤纤维含量分别按GB/T 6435-2014、GB/T 6432-2018、GB/T 6433-2006、GB/T 6438-2007、GB/T 6434-2006、GB/T 20806-2006、NY/T 1459-2007的方法测定,所有养分含量都以干物质基础表示。
1.5 数据处理与统计分析谷实类原料及饲粮肉鸭ME的计算公式1(套算法):
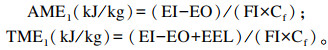
|
谷实类原料及饲粮肉鸭ME的计算公式2(消化率计算法):
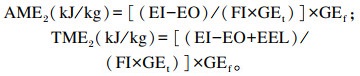
|
式中:AME1和TME1分别为计算公式1(套算法)得出的待测饲料原料的AME(kJ/kg)和TME(kJ/kg);AME2和TME2分别为计算公式2(消化率计算法)得出的待测饲料原料的AME(kJ/kg)和TME(kJ/kg);EI为摄入总能(kJ);EO为排出总能(kJ);FI为采食量(kg);GEt为待测试验饲粮总能(kJ/kg);GEf为待测饲料原料总能(kJ/kg);EEL为内源能量损失(kJ);Cf为待测饲料原料在试验饲粮中的比例(%)。
蛋白质类和其他类原料肉鸭ME的计算公式1(套算法)参考Meloche等[1],具体如下:
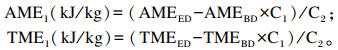
|
蛋白质类和其他类原料肉鸭ME的计算公式2(消化率计算法)参考Woyengo等[5],具体如下:
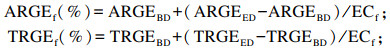
|
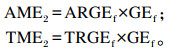
|
式中:AME1和TME1分别为差量法计算公式1(套算法)得出的待测饲料原料的AME(kJ/kg)和TME(kJ/kg);AME2和TME2分别为计算公式2(消化率计算法)得出的待测饲料原料的AME(kJ/kg)和TME(kJ/kg);AMEED和TMEED分别为试验饲粮的AME(kJ/kg)和TME(kJ/kg);AMEBD和TMEBD分别为基础饲粮的AME(kJ/kg)和TME(kJ/kg);C1为试验饲粮中基础饲粮的比例(%);C2为试验饲粮中待测饲料原料的比例(%);ARGEf和TRGEf分别为待测饲料原料的能量表观消化率和真消化率(%);ARGEBD和TRGEBD分别为基础饲粮的能量表观消化率和真消化率(%);ARGEED和TRGEED分别为试验饲粮的能量表观消化率和真消化率(%);GEf为待测饲料原料的总能(kJ/kg);ECf为待测饲料原料在试验饲粮总能中的供能比例(%)。
以SAS 9.0的MEANS模块对基本统计量进行分析。利用SAS 9.0中的PROC GLM模块对肉鸭内源干物质排泄量、有效能值进行方差分析,其统计模型为:Yij=μi+αi+εij,平均值间差异的显著性采用Duncan氏法进行多重比较。饲料原料AME1与AME2或TME1与TME2间的差异采用TTEST模块进行分析。在饲料原料间总能及ME可加性的检验上,通过TTEST模块的Paired选项进行配对t检验。以REG模块通过TEST选项分析计算值对实测值线性回归的斜率与截距分别与1和0的显著性差异,以此来判断实测值与计算值是否相等,从而检验是否具有可加性。
2 结果与分析 2.1 排空-绝食条件下肉鸭内源能损失的变异5个测定批次试验鸭体重及内源能损失的变异见表 4。在肉鸭内源性粪能的测定过程中,在体重的差异上,试验鸭的测试前体重、测试后体重在5个批次间均无显著差异(P>0.05),但在测试过程中体重的损失上,批次间有显著差异(P < 0.05)。在内源粪排泄量的差异上,1~5批次36 h内源粪排泄量在3.01~4.91 g DM/只,其中1~3批间差异不显著(P>0.05),4~5批次间差异不显著(P>0.05),但1~3批次显著低于4~5批次(P < 0.05)。在内源粪能值的差异上,1~5批内源粪能值在12 585~13 083 kJ/kg DM,批次间差异不显著(P>0.05)。36 h内源能损失与36 h内源粪排泄量有类似的规律,1~3批次显著低于4~5批次(P < 0.05)。在数据的变异上,测试前体重、测试后体重、内源粪能值的批内变异系数都在7%以下,而体重损失的批内变异系数在5.39%~18.07%,36 h内源粪排泄量的批内变异系数在8.21%~12.20%,36 h内源能损失的变异系数在9.37%~15.35%。
|
|
表 4 5个批次试验鸭体重及内源能损失的变异 Table 4 Variation of body weight and endogenous energy losses among 5 batches of assay |
由表 5可知,由10种肉鸭饲料原料配制的试验饲粮的GE的实测值与计算值较为接近,最大相差175 kJ/kg。玉米的AME和TME及棉籽粕的AME和TME在2种计算方式上均有显著差异(P < 0.05)。从数值上看,虽然统计检验不显著,但菜籽饼的AME1与AME2(相差242 kJ/kg)和TME1与TME2(相差254 kJ/kg)及玉米DDGS的AME1与AME2(相差459 kJ/kg)和TME1与TME2(相差480 kJ/kg)也有较大的差异。
|
|
表 5 2种公式计算的待测饲料原料ME的差异(干物质基础) Table 5 Difference in ME calculated with 2 formulas for test feed ingredients (DM basis) |
10个验证饲粮GE的实测值与计算值相近,最大相差247 kJ/kg(表 3)。在10个饲粮中原料间ME的可加性(表 6)中,根据公式1计算出的饲料原料AME1获得的饲粮AME计算值与实测值间有显著差异(14 621 kJ/kg vs 14 069 kJ/kg,P=0.000 5),平均值相差552 kJ/kg,相对标准偏差为4.52%。10个饲粮中仅3个饲粮的AME计算值与实测值的差值在400 kJ/kg以内;根据公式2计算出的饲料原料AME2获得的饲粮AME计算值与实测值间有显著差异(14 649 kJ/kg vs 14 069 kJ/kg,P=0.000 1),平均值相差580 kJ/kg,相对标准偏差为4.53%。10个饲粮中3个饲粮的AME实测值与实测值的差值在400 kJ/kg以内;然而,从饲粮AME实测值对计算值1、计算值2的线性回归看,模型的决定系数分别为0.760 3和0.831 1,截距分别为-220 0和-159 1,截距与0均无显著差异(P=0.515 2、P=0.541 9),斜率分别为1.112 7和1.069 0,斜率与1均无显著差异(P=0.623 6、P=0.696 1)。
|
|
表 6 肉鸭饲粮中饲料原料间ME的可加性(干物质基础) Table 6 Additivity of ME among feed ingredients in diets for meat ducks (DM basis) |
 ×100[
×100[ ×100[
×100[根据公式1计算的饲料原料TME1获得的饲粮TME计算值与实测值间无显著差异(15 355 kJ/kg vs 15 216 kJ/kg,P=0.215 3),平均值相差139 kJ/kg,相对标准偏差为2.24%。10个饲粮中7个饲粮的TME实测值与计算值的差值在400 kJ/kg以内;根据公式2计算的饲料原料TME2获得的饲粮TME计算值与实测值间无显著差异(15 386 kJ/kg vs 15 216 kJ/kg,P=0.080 1),平均值相差170 kJ/kg,相对标准偏差为2.03%。10个饲粮中9个饲粮的TME实测值与计算值的差值在400 kJ/kg以内。从饲粮TME实测值对计算值1、计算值2的线性回归看,模型的决定系数分别为0.767 6和0.837 5(P < 0.05),截距分别为-175 4和-106 5,截距与0均无显著差异(P=0.609 8、P=0.685 6),斜率分别为1.105 2和1.058 2,斜率与1均无显著差异(P=0.637 7、P=0.733 1)。
3 讨论 3.1 内源能损失的变异对肉鸭饲料TME的影响采用成年公鸡测定饲料的AME时,AME与采食量间呈负倒数函数关系[11]。由此表明,当采食量过低时,AME的测定值也将偏低。因此,在采用排空强饲法测定饲料的ME时需要扣除包含在排泄物中的内源能损失后计算TME[8]。Dale等[12]通过3次绝食代谢试验测定的成年公鸡的内源能损失分别为10.56、11.40和16.20 kcal/24 h(即44、48和68 kJ/24 h),极差为24 kJ/24 h。若按照玉米风干物的总能16 560 kJ/kg、强饲量50 g计算,则内源能损失可引起能量真消化率和TME最大分别相差2.85%和473 kJ/kg。Ren等[13]通过4次代谢试验测定的黄羽肉鸡的内源能损失分别为16.76、17.02、17.24和18.46 kcal/48 h(即70、71和77 kJ/48 h),极差为7 kJ/48 h,则内源能损失可引起能量真消化率和TME最大分别相差0.9%和142 kJ/kg。杨霞等[14]比较了12个批次成年公鸡内源能损失的差异,不同季节间内源能损失存在显著差异,引起的饲料TME的差异为280~360 kJ/kg。上述在鸡上的试验表明,内源能损失的变异引起饲料ME的变化绝大部分在400 kJ/kg以内。内源能损失的变异主要来源于影响鸡个体代谢状态的因素,如季节[14]、环境温度[15]、年龄[16]等,而与体重损失无相关关系。在本研究中,5个测定批次鸭的体重损失、内源能损失的变异系数(≤15.04%)与鸡[13-14]类似。整体上1~3批次内源能损失比较接近,均低于4~5批次,5个批次内源能损失的极差为23 kJ/36 h,变异系数为24.8%。因此,批次间内源能损失的差异可引起玉米的能量真消化率、TME最大分别相差2.53%和351 kJ/kg。根据Purdue大学通过10个重复批次测定的肉鸭内源能损失为12.1~37.0 kcal/54 h(即51~155 kJ/54 h),变异系数达40.7%[9, 17-20],可以推断不同试验间内源能损失的差异可能会引起玉米的能量真消化率、TME最大分别相差7.53%和1 038 kJ/kg。因此,在多个批次测定饲料的TME中,需要同时测定内源能损失。对内源能损失接近的批次可采用其平均值作为计算TME的内源能。此外,在本试验中,虽然10个饲粮(试验二)的AME和TME的实测值对计算值的回归直线均与Y=X重叠,但AME的实测值总体上低于计算值,而TME的实测值与计算值无显著性差异。因此,排空强饲法测定肉鸭饲料TME的可加性优于AME。
3.2 计算方式对肉鸭饲料原料ME的影响在家禽ME的测定中,考虑到适口性及饲料原料在饲粮配方中的限制,对于非谷食类饲料原料通常采用套算法测定饲料原料的ME。其计算是根据试验饲粮中基础饲粮和待测饲料原料的比例及试验饲粮和基础饲粮的ME采用差减法得出[1]。而在套算法测定猪饲料原料的消化能中,其计算是根据试验饲粮中基础饲粮和待测饲料原料对总能贡献的比例及试验饲粮和基础饲粮的能量消化率先算出待测饲料原料的能量消化率,然后通过能量消化率乘以总能获得待测饲料原料的消化能[4-6]。这2个公式的主要差别是:公式1需要用基础饲粮和试验饲粮的总能;而公式2不仅需要用到基础饲粮和试验饲粮的总能,还需要使用基础饲粮中供能饲料的总能、待测饲料原料的总能。当公式1中饲粮的总能测定值与根据饲粮中供能物质的总能及其比例计算的总能相差较大时,就会导致公式1得出的ME与实际值有较大的偏差,而公式2可以减少公式1因饲粮总能测定误差而将误差转入待测饲料原料中的弊端。本试验中AME1与AME2或TME1与TME2的差异与试验饲粮总能实测值与计算值的差值呈显著的线性相关(r=0.91)。这表明饲粮总能测定的误差与待测饲料原料的ME计算值直接相关[11]。10个验证饲粮的结果表明,公式2测出的AME2、TME2与饲粮ME实测值间的线性关系更高,特别是饲料中TME2比TME1的可加性好,提高了测定数据的准确性。因此,采用公式2计算饲料原料的TME值更科学。
4 结论① 对于肉鸭,套算法和消化率计算法得出的玉米和棉籽粕的AME和TME间存在显著差异,但在其他饲料原料上无显著差异。
② TME比AME具有更好的可加性,采用消化率计算法计算饲料原料的TME有利于提高饲料间TME的可加性。
| [1] |
MELOCHE K J, KERR B J, SHURSON G C, et al. Apparent metabolizable energy and prediction equations for reduced-oil corn distillers dried grains with solubles in broiler chicks from 10 to 18 days of age[J]. Poultry Science, 2013, 92(12): 3176-3183. DOI:10.3382/ps.2013-03290 |
| [2] |
张晋源, 王钰明, 杜中原, 等. 不同实验室间测定单胃动物饲料样品总能的一致性及可加性研究[J]. 动物营养学报, 2021, 33(6): 3581-3591. ZHANG J Y, WANG Y M, DU Z Y, et al. Consistency and additivity in gross energy of monogastric animal feed samples between different laboratories[J]. Chinese Journal of Animal Nutrition, 2021, 33(6): 3581-3591 (in Chinese). DOI:10.3969/j.issn.1006-267x.2021.06.060 |
| [3] |
杜中原, 苏艳芳, 陈凯旋, 等. 2种氧弹热量计测定样品总能的差异及其对能量消化率的影响[J]. 动物营养学报, 2021, 33(2): 1128-1136. DU Z Y, SU Y F, CHEN K X, et al. Difference in two types of oxygen bomb calorimeter to determine gross energy and its effects on energy digestibility of samples[J]. Chinese Journal of Animal Nutrition, 2021, 33(2): 1128-1136 (in Chinese). DOI:10.3969/j.issn.1006-267x.2021.02.053 |
| [4] |
ADEOLA O. Digestion and balance techniques in pigs[M]//LEWIS A J, LEE SOUTHERN L. Swine nutrition. Boca Raton: CRC Press, 2000: 881-894.
|
| [5] |
WOYENGO T A, KIARIE E, NYACHOTI C M. Metabolizable energy and standardized ileal digestible amino acid contents of expeller-extracted canola meal fed to broiler chicks[J]. Poultry Science, 2010, 89(6): 1182-1189. DOI:10.3382/ps.2009-00595 |
| [6] |
KONG C, ADEOLA O. Evaluation of amino acid and energy utilization in feedstuff for swine and poultry diets[J]. Asian-Australasian Journal of Animal Sciences, 2014, 27(7): 917-925. DOI:10.5713/ajas.2014.r.02 |
| [7] |
尹玉港, 赵峰, 李辉, 等. 套算法与消化率计算法测定鸭饲粮脂肪代谢能值的比较研究[J]. 动物营养学报, 2011, 23(7): 1176-1184. YIN Y G, ZHAO F, LI H, et al. A comparative study on the metabolizable energy of dietary fats determined by the difference method and digestibility calculation method in ducks[J]. Chinese Journal of Animal Nutrition, 2011, 23(7): 1176-1184 (in Chinese). DOI:10.3969/j.issn.1006-267x.2011.07.016 |
| [8] |
SIBBALD I R. A bioassay for true metabolizable energy in feedingstuffs[J]. Poultry Science, 1976, 55(1): 303-308. DOI:10.3382/ps.0550303 |
| [9] |
ADEOLA O, RAGLAND D, KING D. Feeding and excreta collection techniques in metabolizable energy assays for ducks[J]. Poultry Science, 1997, 76(5): 728-732. DOI:10.1093/ps/76.5.728 |
| [10] |
WANG Y M, YIN L T, ZHANG H, et al. Accuracy of predicting metabolizable energy from in vitro digestible energy determined with a computer-controlled simulated digestion system in feed ingredients for ducks[J]. Animal Nutrition, 2022, 8(1): 52-60. |
| [11] |
WU S B, CHOCT M, PESTI G. Historical flaws in bioassays used to generate metabolizable energy values for poultry feed formulation: a critical review[J]. Poultry Science, 2020, 99(1): 385-406. DOI:10.3382/ps/pez511 |
| [12] |
DALE N M, FULLER H L. Endogenous energy losses of fed versus fasted roosters[J]. Poultry Science, 1982, 61(5): 898-901. DOI:10.3382/ps.0610898 |
| [13] |
REN L Q, TAN H Z, ZHAO F, et al. Using corn starch as basal diet to determine the true metabolizable energy of protein feedstuffs in Chinese yellow chickens[J]. Poultry Science, 2012, 91(6): 1394-1399. DOI:10.3382/ps.2011-01838 |
| [14] |
杨霞, 党方昆, 赵峰, 等. 鸡内源总能的变异及其对饲料原料真代谢能值的影响[J]. 动物营养学报, 2016, 28(7): 2005-2012. YANG X, DANG F K, ZHAO F, et al. Variation of gross endogenous energy and its effect on value of true metabolizable energy of feedstuffs in roosters[J]. Chinese Journal of Animal Nutrition, 2016, 28(7): 2005-2012 (in Chinese). |
| [15] |
TOMONAGA S, OKUYAMA H, TACHIBANA T, et al. Effects of high ambient temperature on plasma metabolomic profiles in chicks[J]. Animal Science Journal, 2018, 89(2): 448-455. DOI:10.1111/asj.12951 |
| [16] |
YANG Z, PIRGOZLIEV V R, ROSE S P, et al. Effect of age on the relationship between metabolizable energy and digestible energy for broiler chickens[J]. Poultry Science, 2020, 99(1): 320-330. DOI:10.3382/ps/pez495 |
| [17] |
KING D, RAGLAND D, ADEOLA O. Apparent and true metabolizable energy values of feedstuffs for ducks[J]. Poultry Science, 1997, 76(10): 1418-1423. DOI:10.1093/ps/76.10.1418 |
| [18] |
HONG D, RAGLAND D, ADEOLA O. Additivity and associative effects of metabolizable energy and amino acid digestibility in barley and canola meal for white pekin ducks[J]. Poultry Science, 2001, 80(11): 1600-1606. DOI:10.1093/ps/80.11.1600 |
| [19] |
HONG D, RAGLAND D, ADEOLA O. Additivity and associative effects of metabolizable energy and amino acid digestibility of corn, soybean meal, and wheat red dog for white Pekin ducks[J]. Journal of Animal Science, 2002, 80(12): 3222-3229. DOI:10.2527/2002.80123222x |
| [20] |
ADEOLA O. Energy values of feed ingredients for white Pekin ducks[J]. International Journal of Poultry Science, 2003, 2(5): 318-323. DOI:10.3923/ijps.2003.318.323 |