




2. 大连海洋大学, 大连 116023;
3. 温州医科大学, 温州 325000
2. Dalian Ocean University, Dalian 116023, China;
3. Wenzhou Medical University, Wenzhou 325000, China
随着世界人口的增长和人民生活水平的提高,人们对水产品的需求量不断增加。近十几年来,水产养殖正逐渐成为提高全球水产品供给量的主要方式,而水产养殖集约化和规模化的生产又依赖于水产饲料工业的发展[1]。由于鱼粉营养全面、适口性好,一直是水产饲料中不可或缺的优质蛋白质源。近年来,随着鱼粉价格的不断上扬,采用植物蛋白质源替代鱼粉逐渐受到研究者和养殖者的重视。棉籽粕是一种资源丰富、蛋白质含量高、价格相对便宜、可以用于水产饲料的优质植物蛋白质源。但是,由于棉籽粕中含有对动物有害的游离棉酚而在使用中受到限制。Yildirim等[2]研究发现,饲料中游离棉酚达到300 mg/kg时,斑点叉尾 幼鱼生长受到抑制,红细胞、血红素和血红蛋白浓度变低。Lim[3]对凡纳滨对虾的研究发现,用26.5%浸提棉籽粕等氮替代40%的复合海洋蛋白质源,对虾的生长、饲料利用和成活率均无显著变化,但替代比例再增加会导致其生长性能降低。Cheng等[4]对虹鳟幼鱼的研究发现,当棉籽粕在饲料中添加量超过15%时,鱼体增重率和饲料转化效率显著降低。以上研究表明,当饲料中少量添加棉籽粕时,不会抑制水生动物的生长;而添加量较高时,则会显著降低饲料的适口性,改变饲料的氨基酸组成,从而抑制水生动物生长。由此可见,用棉籽粕部分替代其他蛋白质源是可行的。如果对棉籽粕进行微生物脱毒处理,利用微生物在发酵过程中对游离棉酚的转化降解作用,则可改善棉籽粕的蛋白质含量以及适口性,从而进一步提高棉籽粕在饲料中的利用率。
中华绒螯蟹(Eriocheir sinensis),俗称河蟹、大闸蟹,是我国重要的淡水养殖珍品,由于其营养丰富、味道鲜美,倍受国内外市场青睐。关于其配合饲料中植物蛋白质源替代鱼粉的研究已有一些报 道[5,6,7,8,9,10],而有关发酵棉籽粕(FCSM)在中华绒螯 蟹配合饲料中的应用还未见报道。本试验拟通过在配合饲料中添加普通棉籽粕(CSM)和发酵棉籽粕来部分或全部替代鱼粉饲喂中华绒螯蟹幼蟹,旨在研究不同替代水平对中华绒螯蟹幼蟹生长性能、体成分及肝胰腺消化酶活性的影响,并探讨普通棉籽粕和发酵棉籽粕作为鱼粉替代蛋白质源时在饲料中的适宜添加量,以期充分利用植物蛋白质源,为中华绒螯蟹饲料配方中合理添加棉籽粕提供相关的参考。
本试验采用自主筛选的棉酚降解菌种(HQ-1)[11]对普通棉籽粕进行固体发酵,30 ℃下发酵72 h后收集发酵棉籽粕,60 ℃烘干,用万能粉碎机将其粉碎,过60目筛,-20 ℃冰箱中保存备用。发酵好的棉籽粕颜色略深于普通棉籽粕,无异味,无霉变结块。普通棉籽粕与发酵棉籽粕的化学成分见表1。
| 表1 普通棉籽粕与发酵棉籽粕的化学成分(风干基础) Table 1 Chemical composition of CSM and FCSM (air-dry basis) % |
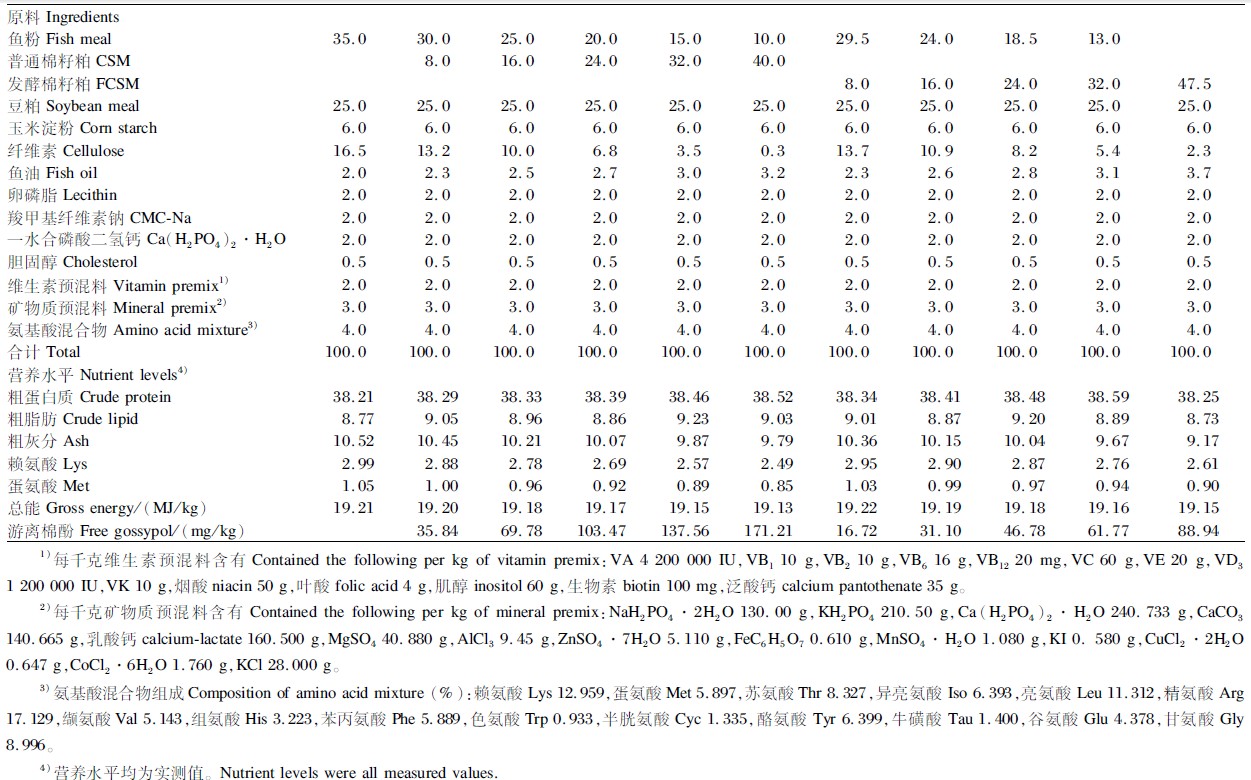
以秘鲁红鱼粉、豆粕、发酵棉籽粕或普通棉籽粕为蛋白质源,鱼油、豆油、卵磷脂为脂肪源配制成11组等氮(粗蛋白质,38.39%)等能(总能,19.18 MJ/kg)的试验饲料。普通棉籽粕添加量设为0、8.0%、16.0%、24.0%、32.0%、40.0%(分别记为CSM0、CSM8、CSM16、CSM24、CSM32、CSM40组);发酵棉籽粕添加量设为8.0%、16.0%、24.0%、32.0%、47.5%(分别记为FCSM8、FCSM16、FCSM24、FCSM32、FCSM47.5组)。以CSM0组为对照组,试验饲料组成及营养水平见表2,试验饲料氨基酸组成见表3。
| 表2 试验饲料组成及营养水平(风干基础) Table 2 Composition and nutrient levels of experimental diets (air-dry basis) % |
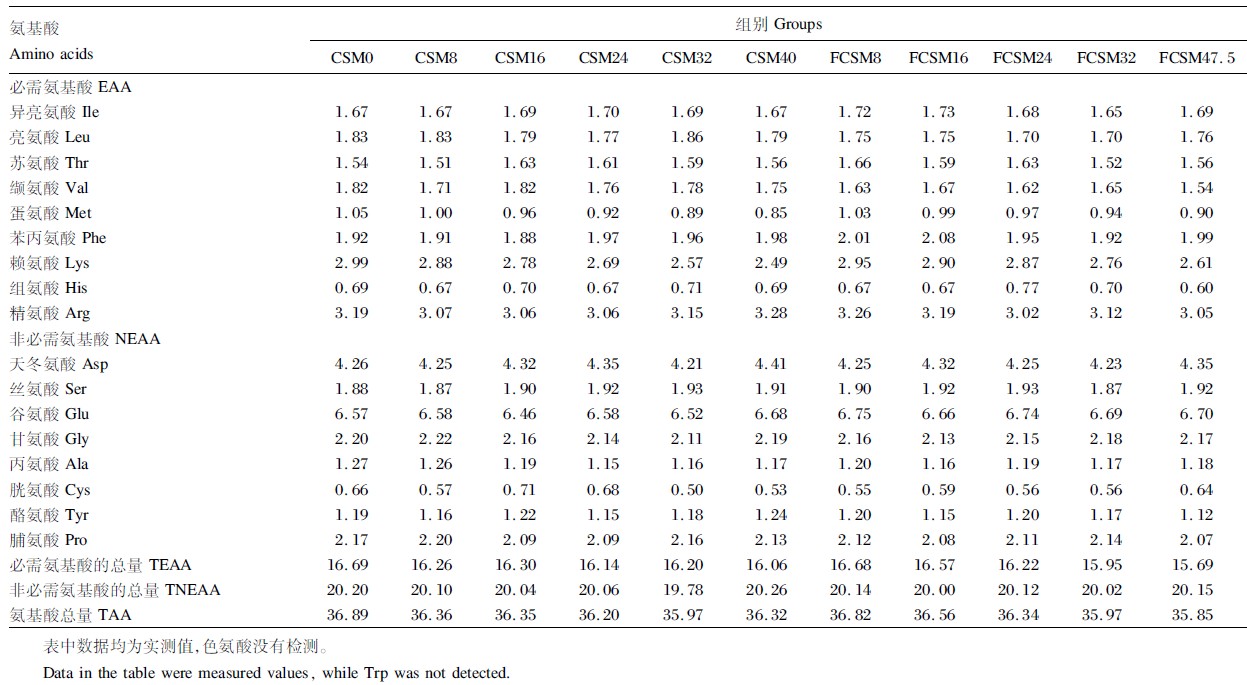
| 表3 试验饲料氨基酸组成 Table 3 Composition of amino acids in experimental diets % |
试验饲料制作时,将所有原料粉碎并通过60目筛,采用逐级扩大法将维生素和矿物质预混料等微量成分按比例充分混匀,然后加入鱼油、卵磷脂和水搅拌混匀,通过饲料机(型号SLX-80,上海亚励科技贸易有限公司)挤压制成直径为1.5 mm的长条,55 ℃烘干,自然冷却后破碎成粒径为1~2 mm的破碎料,置于4 ℃冰箱内密封保存备用。
试验用中华绒螯蟹幼蟹购自上海市崇明岛长江口中华绒螯蟹养殖基地,采用经过沉淀、过滤后的天然池塘水进行养殖。幼蟹于水泥池中暂养1周后,挑取体重均一、附肢完整、身体健壮的个体作为试验对象,随机分成11个组,每组3个重复,以重复为单位放入300 L的水箱中,每箱20只。
水箱中放置足够数量的网片和聚氯乙烯(PVC)管作为隐蔽物,箱内水深约30 cm。试验幼蟹的初始体重为(0.56±0.04) g,每组投喂1种试验饲料。养殖期间,采用饱食投喂法,每天08:00和16:30各投喂1次,次日吸污并补充养殖用水,试验期为6周。每周测量1次水质,水温为(28±2) ℃、溶解氧>5 mg/L、pH 7.5±0.5,连续充气增氧。
养殖试验结束后,停食24 h排空肠道,分组称重,并统计成活数。每个养殖箱中随机选取成活数的1/2立即烘干,制成全蟹样品用于体成分分析,另外1/2立即解剖,取其肝胰腺用于消化酶活性测定,-80 ℃保存备用。
生长性能指标参照以下公式计算:
成活率(SR,%)=(终末蟹数/初始蟹数)×100;
增重率(WGR,%)=[(终末体重-初始体重)/初始体重]×100;
特定生长率(SGR,%/d)=[(ln终末体重-ln初始体重)/试验天数]×100;
蛋白质效率(PER,%)=[(终末体重-初始体重)/蛋白质摄入量]×100;
饲料系数(FCR)=投喂量/(终末体重-初始体重)。
饲料原料及全蟹样品中水分含量采用105 ℃恒重法测定,粗蛋白质含量采用凯氏定氮法测定,粗脂肪含量采用乙醚抽提法测定,粗灰分含量采用马福炉550 ℃灼烧法测定,游离棉酚含量按照《饲料中游离棉酚的测定方法》(GB/T 13086—1991)中方法测定。
粗酶液制备:用分析天平准确称量肝胰腺,按组织重量加入9倍体积的预冷0.9%生理盐水,制成10%的组织匀浆,4 ℃、2 500 r/min离心10 min,取上清液待测。采用南京建成生物工程研究所生产的试剂盒测定胰蛋白酶、胃蛋白酶、淀粉酶和脂肪酶活性,蛋白质含量采用考马斯亮蓝法测定。胰蛋白酶活性定义为:在pH 8.0、37 ℃条件下,每毫克组织蛋白质中含有的胰蛋白酶每分钟使吸光度变化0.003为1个酶活性单位(U/mg prot)。胃蛋白酶活性定义为:每毫克组织蛋白质在37 ℃条件下每分钟分解蛋白质生成1 μg氨基酸为1个酶活性单位(U/mg prot)。淀粉酶活性定义为:每毫克组织蛋白质在37 ℃条件下与底物作用30 min,水解10 mg淀粉为1个酶活性单位(U/mg prot)。脂肪酶活性定义为:在37 ℃条件下,每克组织蛋白质在本反应体系中与底物反应1 min,每消耗1 μmol底物为1个酶活性单位(U/g prot)。纤维素酶活性采用酶联免疫吸附测定(ELISA)试剂盒测定,试剂盒由南京建成生物工程研究所生产。
试验数据采用平均值±标准差(mean±SD)表示,采用SPSS 20.0中的one-way ANOVA软件包对所得数据进行单因素方差分析,若有显著差异(P<0.05),再用Duncan氏法进行多重比较。
由表4可知,CSM8、CSM16、FCSM8、FCSM16、FCSM24组的增重率、特定生长率、成活率与对照组差异不显著(P>0.05),CSM40组的增重率、特定生长率、成活率显著低于其他各组(P<0.05),且蛋白质效率达最小值,饲料系数达最大值。与对照组相比,FCSM8组的蛋白质效率无显著差异(P>0.05),其他各组均显著降低(P<0.05)。在相同替代水平下,发酵棉籽粕组的各项生长性能指标均好于普通棉籽粕组,完全替代鱼粉组(FCSM47.5组)的增重率、特定生长率和成活率均显著高于CSM40组(P<0.05)。
| 表4 普通棉籽粕和发酵棉籽粕替代鱼粉对中华绒螯蟹生长性能的影响 Table 4 Effects of fish meal replacement by CSM and FCSM on growth performance of juvenile Chinese mitten crab (Eriocheir sinensis) |
以饲料中游离棉酚含量为横坐标,特定生长率为纵坐标,通过二次曲线回归分析得出,当普通棉籽粕饲料中的游离棉酚含量不超过62.18 mg/kg时,中华绒螯蟹幼蟹的特定生长率不会明显降低(图 1)。同时,当发酵棉籽粕饲料中的游离棉酚含量不超过44.36 mg/kg时,中华绒螯蟹幼蟹的特定生长率不会明显降低(图 2)。
 | 图 1 中华绒螯蟹幼蟹特定生长率与普通棉籽粕饲料中游离棉酚含量的关系 Fig.1 Relationship between the content of free gossypol in CSM diets and specific growth rate of juvenile Chinese mitten crab (Eriocheir sinensis) |
 | 图 2 中华绒螯蟹幼蟹特定生长率与发酵棉籽粕饲料中游离棉酚含量的关系 Fig. 2 Relationship between the content of free gossypol in FCSM diets and specific growth rate of juvenile Chinese mitten crab (Eriocheir sinensis) |
由表5可知,CSM8、FCSM8、FCSM16和FCSM24组的全蟹粗蛋白质含量与对照组差异不显著(P>0.05),其他组则均显著低于对照组(P<0.05)。在普通棉籽粕组中,随替代水平的升高,全蟹粗蛋白质含量显著下降(P<0.05),CSM40组的粗蛋白质含量仅为11.40%;在发酵棉籽粕组中也呈类似的趋势,FCSM47.5组的全蟹粗蛋白质含量为11.75%,显著高于CSM40组(P<0.05)。不同水平的普通棉籽粕和发酵棉籽粕替代鱼粉对中华绒螯蟹幼蟹全蟹水分、粗脂肪和粗灰分含量均无显著影响(P>0.05)。
| 表5 普通棉籽粕和发酵棉籽粕替代鱼粉对中华绒螯蟹全蟹体成分的影响
Table 5 Effects of fish meal replacement by CSM and FCSM on body composition of juvenile Chinese mitten crab (Eriocheir sinensis) %
|
由表6可知,FCSM8组的肝胰腺胃蛋白酶活性略低于对照组,但差异不显著(P>0.05),而其他各组的肝胰腺胃蛋白酶活性均显著低于对照组(P<0.05),且以CSM40组最低,仅为8.28 U/mg prot。随着普通棉籽粕或发酵棉籽粕替代水平的升高,肝胰腺胃蛋白酶活性呈下降趋势,但相同替代水平下,发酵棉籽粕组的肝胰腺胃蛋白酶活性要高于普通棉籽粕组。肝胰腺胰蛋白酶活性与胃蛋白酶活性的变化趋势相似,除对照组外,以FCSM8组的肝胰腺胰蛋白酶活性最高,为1 099.84 U/mg prot,而CSM40组的胰蛋白酶活性则最低,为497.47 U/mg prot,显著低于其他各组(P<0.05)。
| 表6 普通棉籽粕和发酵棉籽粕替代鱼粉对中华绒螯蟹幼蟹肝胰腺消化酶活性的影响
Table 6 Effects of fish meal replacement by CSM and FCSM on digestive enzyme activities in hepatopancreas of juvenile Chinese mitten crab (Eriocheir sinensis)
|
不同水平的发酵棉籽粕和普通棉籽粕替代鱼粉对中华绒螯蟹幼蟹肝胰腺脂肪酶活性的影响差异不显著(P>0.05),且各组脂肪酶活性均较低,平均为40.53 U/g prot。
对照组的肝胰腺淀粉酶活性最低,为5.96 U/mg prot,显著低于其他各组(P<0.05)。在普通棉籽粕组中,肝胰腺淀粉酶活性随替代水平的升高而逐渐升高,在发酵棉籽粕组中也呈现出类似的趋势。其中,以CSM40组的肝胰腺淀粉酶活性最高,为9.39 U/mg prot。肝胰腺纤维素酶活性的变化趋势与淀粉酶活性类似,以对照组的肝胰腺纤维素酶活性最低,为7.82 μg/mL,显著低于其他各组(P<0.05)。在相同替代水平下,普通棉籽粕组的肝胰腺纤维素酶活性要显著高于发酵棉籽粕组(P<0.05)。其中,CSM40组的肝胰腺纤维素酶活性达最大值30.63 μg/mL。
研究发现,在以植物蛋白质源作为饲料中唯一蛋白质源时,大多数水生动物的生长都将受到抑制。但是,当植物蛋白质源的添加量控制在一定范围时,水生动物的生长性能不会出现较大差异,当添加量超过该范围时,其生长性急剧下降[9]。本试验的结果也验证了这一点,即随着普通棉籽粕或发酵棉籽粕替代鱼粉水平的升高,中华绒螯蟹幼蟹的增重率、特定生长率、蛋白质效率和成活率均呈下降趋势。其中,CSM8和CSM16组的增重率、特定生长率和成活率与对照组差异不显著,但是,当普通棉籽粕添加量为40%时(CSM40组),中华绒螯蟹幼蟹的成活率低于50%,仅为48.33%。这是本试验提前结束,只进行了6周养殖试验的重要原因之一。上述结果说明中华绒螯蟹幼蟹对棉籽粕中的有害物质——游离棉酚具有一定的耐受力,但必须控制添加量。由此得出,本试验中普通棉籽粕的适宜添加量为16%。此外,回归模型分析发现,当普通棉籽粕饲料中游离棉酚含量不超过62.18 mg/kg时,对中华绒螯蟹幼蟹的生长没有明显的抑制作用。另外,有研究者提出,高水平的植物蛋白质源替代鱼粉除了其中抗营养因子的毒性对水生动物生长产生影响外,还可能由于植物蛋白质源中氨基酸组成不能满足水生动物生长所需的某些必需氨基酸(尤其是赖氨酸和蛋氨酸)而对生长产生抑制作用,所以需在饲料中补充晶体氨基酸[9]。从棉籽粕的氨基酸组成来看,除精氨酸、苯丙氨酸含量较多外,其他氨基酸的含量均低于鱼和虾、蟹的生长需要[15]。孙立梅等[9]研究了在高水平棉籽粕饲料中补充蛋氨酸对中华绒螯蟹幼蟹[(0.39±0.02) g]生长的影响,结果表明,添加0.42%蛋氨酸的40%棉籽粕饲料组的增重率、特定生长率和饲料系数与全鱼粉组相比均无显著差异,说明在高水平棉籽粕饲料中适当补充外源蛋氨酸可以明显改善饲料的适口性,有效提高中华绒螯蟹幼蟹的生长性能。
目前,关于中华绒螯蟹饲料蛋白质源替代的研究主要集中在采用普通植物蛋白质(豆粕、棉籽粕、菜籽粕、花生粕等)替代鱼粉[5-10,12-14],关于发酵棉籽粕在中华绒螯蟹饲料中的应用还未见报道。而棉籽粕经微生物发酵或脱酚处理后,其营养价值和适口性均明显得到改善,利用率大大提高,这已经在许多水产动物中得到了验证[16,17,18,19,20,21,22]。孔丽等[16]在研究发酵棉籽粕对异育银鲫生长性能的影响时发现,与对照组(不添加发酵棉籽粕组)相比,6%发酵棉籽粕组的增重率和特定生长率显著增加。吴秀峰等[17]研究了用脱酚棉籽粉(SECM)替代0、25%和40%的鱼粉对西伯利亚鲟幼鱼(57.36 g)生长性能的影响,结果显示,SECM 25C组(替代25%且晶体氨基酸平衡组)的增重率、特定生长率和蛋白质效率显著高于SECM 25B组(替代25%且血粉氨基酸平衡组);SECM 25C组的饲料系数与对照组差异不显著,其他试验组均显著高于对照组。薛敏等[18]以0.17 g左右的南美白对虾虾苗为试验对象,用脱酚棉籽蛋白替代0、20%、35%、50%的鱼粉蛋白(对照组鱼粉添加量为36%),结果显示,20%替代组南美白对虾的增重率、特定生长率、饲料系数和蛋白质效率与对照组相比无显著差异。这与本试验的研究结果相似,本试验中FCSM8、FCSM16和FCSM24组的增重率、特定生长率和成活率与对照组均无显著差异,因此,建议发酵棉籽粕的适宜添加量为24%。可见,由于加工工艺和试验对象不同,发酵棉籽粕的适宜添加量差异较大。此外,回归模型分析发现,发酵棉籽粕中游离棉酚含量不超过44.36 mg/kg时,对中华绒螯蟹幼蟹生长没有明显的抑制作用。一般认为,鲑科鱼类饲料中游离棉酚的含量应限制在100 mg/kg以下[23]。而凡纳滨对虾在摄食含1 100 mg/kg游离棉酚饲料时,其生长才受到抑制[3]。这与本试验的结果差异较大,有待进一步证实,但也说明不同种类的水产动物对游离棉酚的敏感程度不同,而中华绒螯蟹幼蟹对游离棉酚的毒性更敏感。此外,CSM16组与FCSM24组相比,增重率、特定生长率、蛋白质效率、成活率和饲料系数均无显著差异,说明添加发酵棉籽粕可以更多地减少鱼粉用量,降低饲料成本。同时,在相同替代水平下,发酵棉籽粕的效果要优于普通棉籽粕,其原因可能有2方面:一是普通棉籽粕经过微生物发酵后,游离棉酚等抗营养因子明显减少,使得饲料的适口性得到改善,动物的采食量增加;二是微生物脱毒处理的整个过程都比较温和,不会破坏其原有的营养成分,微生物在脱毒的同时还能够生成菌体蛋白和维生素,使棉籽饼粕的营养价值和食用价值均得到提高(表1和表3)。
在体成分方面,陈亮[5]的研究表明,不同水平棉籽粕替代鱼粉对中华绒螯蟹幼蟹全蟹水分含量没有显著影响;对照组全蟹粗灰分含量显著低于66%棉籽粕添加组,而全蟹粗蛋白质含量则高于各棉籽粕添加组,但仅与66%棉籽粕添加组之间存在显著差异。薛敏等[18]研究认为,不同水平脱酚棉籽蛋白替代鱼粉蛋白对南美白对虾全虾粗蛋白质、粗脂肪、粗灰分和水分含量的影响均不显著。伍代勇等[23]认为,不同水平棉籽粕替代鱼粉对凡纳滨对虾肌肉成分无显著影响。Yue等[24]和El-Saidy等[25]研究均表明,不同水平棉籽粕替代鱼粉对尼罗罗非鱼肌肉成分无显著影响。以上结果与本试验结果略有不同,本试验结果显示,不同水平棉籽粕和发酵棉籽粕替代鱼粉对中华绒螯蟹全蟹水分、粗脂肪和粗灰分含量均无显著影响,但随着普通棉籽粕或发酵棉籽粕替代水平的升高,全蟹粗蛋白质含量逐渐减少,且CSM16、CSM24、CSM32、CSM40、FCSM32和FCSM47.5组与对照组相比显著下降。这可能与试验动物种类、替代水平和替代方法等不同有关。
研究表明,甲壳动物幼体消化酶活性的高低直接反映了其对营养物质的消化吸收能力,并且与营养物质含量呈正相关。肝胰腺是甲壳动物主要的消化腺,其消化酶活性远远超过胃和肠道。本试验结果表明,华绒螯蟹幼蟹肝胰腺蛋白酶活性较高,且胰蛋白酶活性高于胃蛋白酶活性,这与其他学者对甲壳动物消化酶活性的研究结果[26,27,28,29]一致。本研究还发现,随着饲料中普通棉籽粕或发酵棉籽粕替代水平的升高,胰蛋白酶和胃蛋白酶活性均显著下降,而且在相同替代水平下,发酵棉籽粕组高于普通棉籽粕组。棉籽粕对中华绒螯蟹肝胰腺蛋白酶活性产生影响,其主要原因是现阶段棉籽粕的加工普遍采用的是高温蒸炒压榨工艺,温度过高,破坏了赖氨酸、蛋氨酸及其他必需氨基酸。此外,普通棉籽粕中的游离棉酚会与赖氨酸的氨基结合发生褐变反应,从而大大降低赖氨酸的有效成分[30],机体不能够获得足够的、平衡的氨基酸,蛋白酶也就不能顺利合成和分泌,最终会影响饲料营养物质的消化和利用[5]。经过微生物发酵处理后,发酵棉籽粕中游离棉酚的含量大大降低,氨基酸组成也趋于合理,使得蛋白酶合成和分泌增加。
中华绒螯蟹属于杂食性甲壳动物,其肝胰腺中脂肪酶活性与动物性饲料有关,而淀粉酶和纤维素酶活性则与植物性饲料有关。本试验显示,中华绒螯蟹幼蟹肝胰腺脂肪酶活性各组间无显著差异,并且各组的活性均较低,这与其他学者关于甲壳动物脂肪酶活性的研究结果[28,29]一致。碳水化合物是生物体内三大能源物质之一,目前关于甲壳动物淀粉酶和纤维素酶活性的研究存在较大争议。Hirche等[31]认为,甲壳动物幼体相对较高的淀粉酶活性并不是对食物中相应生化成分的反映,而可能是祖先物种食草性幼体系统发育的残余。关于甲壳动物是否能分泌纤维素酶尚无定论。于书坤等[32]认为,在滤食性和杂食性动物中纤维素酶完全由肠道内微生物产生,而在肉食性甲壳动物中纤维素酶是退化的痕迹,毫无功能。本试验结果显示,饲料中添加普通棉籽粕和发酵棉籽粕均能够提高中华绒螯蟹幼蟹肝胰腺淀粉酶和纤维素酶的活性,而棉籽粕中碳水化合物(28.69%)和粗纤维含量(10.24%)都远远高于鱼粉(分别为8.00%和0.50%),这说明中华绒螯蟹幼蟹肝胰腺对淀粉和纤维素的消化能力与食物组成有关。本试验还发现,在相同替代水平下,普通棉籽粕组的肝胰腺纤维素酶活性高于发酵棉籽粕组,这可能是由于本试验中所用的发酵棉籽粕是由真菌发酵普通棉籽粕所得,而真菌本身能够分泌纤维素酶,导致发酵棉籽粕中的粗纤维含量(4.47%)远远低于普通棉籽粕(10.24%)。中华绒螯蟹幼蟹肝胰腺中高活性的淀粉酶和纤维素酶无论是自身分泌的还是肠道细菌产生的,只要它摄取了含淀粉和纤维素的食物, 这2种酶就能够起作用。因此,在中华绒螯蟹幼蟹配合饲料中添加含淀粉和纤维素较高的植物蛋白质原料是可行的。
① 作为中华绒螯蟹幼蟹饲料中鱼粉的替代蛋白质源时,普通棉籽粕的适宜添加量为16%,发酵棉籽粕的适宜添加量为24%。
② 在相同替代水平下,发酵棉籽粕的饲喂效果比普通棉籽粕好。
| [1] | 周歧存, 麦康森, 刘永坚, 等.动植物蛋白源替代鱼粉研究进展[J]. 水产学报, 2005, 29(3):404-410. ( 1) 1)
|
| [2] | YILDIRIM M, LIM C, WAN P J, et al.Growth performance and immune response of channel catfish (Ictalurus punctatus) fed diets containing graded levels of gossypol-acetic acid[J]. Aquaculture, 2003, 219(1/2/3/4):75l-768. (1)
|
| [3] | LIM C.Substitution of cottonseed meal for marine animal protein in diets for Penaeus vannamei[J]. Journal of the World Aquaculture Society, 1996, 27(4):402-409. (2)
|
| [4] | CHENG Z J, HARDY R W.Apparent digestibility coefficients and nutritional value of cottonseed meal for rainbow trout (Oncorhynchus mykiss)[J]. Aquaculture, 2002, 212(1/2/3/4):361-372. (1)
|
| [5] | 陈亮.不同棉籽粕水平饲料对中华绒螯蟹幼蟹生长和相关生理生化指标的影响[D]. 硕士学位论文.上海:华东师范大学, 2011. (3)
|
| [6] | 陈立侨, 堵南山, 赖伟.中华绒螯蟹蟹种配饵中豆饼替代部分鱼粉的适宜含量[J]. 水产学报, 1994, 18(1):24-31. (1)
|
| [7] | JIANG H B, CHEN L Q, LI E C, et al.Partial or total replacement of soybean meal by cottonseed meal in practical diets for Chinese mitten crab, Eriocheir sinensis:effects on oxygen consumption, ammonia excretion, O∶ N ratio and amino transferases activities[J].Turkish Journal of Fisheries and Aquatic Sciences, 2012, 12(3):547-554. (1)
|
| [8] | JIANG H B, SUN L Y, CHEN L Q, et al.Partial or total replacement of soybean meal by cottonseed meal in practical diets for Chinese mitten crab (Eriocheir sinensis):effects on growth, feed utilization, and body composition[J]. Israeli Journal of Aquaculture-Bamigdeh, 2012, 64:754-760. (1)
|
| [9] | 孙立梅, 陈立侨, 李二超, 等.高比例棉粕饲料中补充蛋氨酸对中华绒螯蟹幼蟹摄食、生长及抗氧化酶活性的影响[J]. 水生生物学报, 2013, 37(2):336-343. (4)
|
| [10] | JIANG H B, CHEN L Q, QIN J G, et al.Partial or complete substitution of fish meal with soybean meal and cottonseed meal in Chinese mitten crab Eriocheir sinensis diets[J]. Aquaculture International, 2013, 21(3):617-628. (1)
|
| [11] | YANG X, GUO J L, SUN J Y.Biodegradation of free-gossypol by a new fungus isolated from cotton planted soil[J]. African Journal of Microbiology Research, 2011, 5(19):3066-3072. (1)
|
| [12] | 陈权文.中华绒螯蟹对常用饲料蛋白源利用性研究[D]. 硕士学位论文.苏州:苏州大学, 2009. (0)
|
| [13] | 张璐, 陈立侨, 洪美玲, 等.中华绒螯蟹对11种饲料原料蛋白质和氨基酸的表观消化率[J]. 水产学报, 2007, 31(1):116-121. (0)
|
| [14] | LI M H, ROBINSON E H.Use of cottonseed meal in aquatic animal diets:a review[J]. North American Journal of Aquaculture, 2006, 68(1):14-22. (0)
|
| [15] | ROBINSON E H, LI M H.Use of plant proteins in catfish feeds:replacement of soybean meal with cottonseed meal and replacement of fish meal with soybean meal and cottonseed meal[J]. Journal of the World Aquaculture Society, 1994, 25(2):271-276. (1)
|
| [16] | 孔丽, 张伟, 叶元土, 等.氨基酸粉·发酵棉粕和发酵菜粕在异育银鲫饲料中的应用效果[J]. 安徽农业科学, 2011, 39(6):3610-3612, 3623. (1)
|
| [17] | 吴秀峰, 薛敏, 郭利亚, 等.脱酚棉籽粉替代部分鱼粉对西伯利亚鲟幼鱼生长、体成分及血清生化指标的影响[J]. 动物营养学报, 2010, 22(1):117-124. (2)
|
| [18] | 薛敏, 吴秀峰, 郭利亚, 等.脱酚棉籽蛋白在水产饲料中的应用[J]. 中国畜牧杂志, 2007, 43(8):55-58. (3)
|
| [19] | 罗琳, 薛敏, 吴秀峰, 等.脱酚棉籽蛋白对日本鲈的生长、体成分及营养成分表观消化率的影响[J]. 水产学报, 2005, 29(6):866-870. (1)
|
| [20] | 林仕梅, 毛述宏, 关勇, 等.罗非鱼低鱼粉饲料中脱酚棉籽蛋白替代鱼粉的研究[J]. 动物营养学报, 2011, 23(12):2331-2338. (1)
|
| [21] | KONG L, ZHANG W, YE Y T, et al.Application effects of amino acid powder, fermented rapeseed meal and fermented cottonseed meal in feeds for allogynogenetic crucian carp[J].Animal Husbandry and Feed Science, 2010, 2(6):7-10, 13. (1)
|
| [22] | 陈道仁, 冷向军, 肖昌武.发酵棉粕对草鱼生长的影响[J]. 中国饲料添加剂, 2010(6):34-36. (1)
|
| [23] | 伍代勇, 叶元土, 张宝彤, 等.棉籽粕和菜籽粕对凡纳滨对虾生长、非特异免疫和体成分的影响[J]. 中国饲料, 2009(23):27-30. (2)
|
| [24] | YUE Y R, ZHOU Q C.Effect of replacing soybean meal with cottonseed meal on growth, feed utilization, and hematological indexes for juvenile hybrid tilapia, Oreochromis niloticus×O.aureus[J]. Aquaculture, 2008, 284(1/2/3/4):185-189. (1)
|
| [25] | EI-SAIDY D M S D, GABER M M.Use of cottonseed meal supplemented with iron for detoxification of gossypol as a replacement of fish meal in Nile tilapia, Oreochromis niloticus (L.) diets[J]. Aquaculture Research, 2004, 35(9):859-865. (1)
|
| [26] | 潘鲁青, 王克行.中华绒螯蟹幼体消化酶活力与氨基酸组成的研究[J]. 中国水产科学, 1997, 4(2):13-20. (1)
|
| [27] | 潘鲁青, 王奎琪.三疣梭子蟹幼体消化酶活力及氨基酸组成的研究[J]. 水产学报, 1997, 21(3):246-251. (1)
|
| [28] | 汤鸿, 李少菁, 王桂忠, 等.锯缘青蟹幼体消化酶活力[J]. 厦门大学学报:自然科学版, 1995, 34(1):89-93. (2)
|
| [29] | 潘鲁青.四种虾蟹类幼体消化酶活力的比较研究[J]. 青岛海洋大学学报, 1997, 27(3):313-318. (2)
|
| [30] | 萧培珍, 叶元土, 张宝彤, 等.棉籽粕的营养价值及其在水产饲料上的应用[J]. 饲料工业, 2009, 30(18):49-5l. (1)
|
| [31] | HIRCHE H J, ANGER K.Digestive enzyme activities during larval development of Hyasaraneus (Decapoda, Majidae)[J]. Comparative Biochemistry and Physiology Part B:Comparative Biochemistry, 1987, 87(2):297-302. (1)
|
| [32] | 于书坤, 张树荣.虾类及甲壳动物消化酶研究的现状[J]. 海洋科学, 1986, 10(2):60-63. (1)
|